Н и НН в прилагательных
В прилагательных пишется как одна буква -Н-, так и -НН-. Правильный выбор сделаем, определив, от слова какой части речи и с помощью каких суффиксов образуются прилагательные.
Для того чтобы выбрать одну и две буквы -Н- в написании имен прилагательных, определим, с помощью каких суффиксов они образованы от существительных или глаголов несовершенного и совершенного вида.
Написание -Н- и -НН- в прилагательных, образованных от существительных
Если прилагательное образовано от существительного с помощью суффиксов -ан-, -ян-, -ин-, то эти суффиксы пишутся с одной буквой -Н-, например:
- песок — песчаный пляж;
- рожь — ржаной хлеб;
- береста — берестяное лукошко;
- серебро — серебряный крестик;
- осёл — ослиный крик;
- соловей — соловьиное пение.


Обратим внимание, что коренное «н» пишется в прилагательных:
Запомним написание слов-исключений:
- олово — оловянный
- дерево — деревянный
- стекло — стеклянный
Прилагательные образуются от существительных с помощью суффиксов -онн- и -енн-, например:
- лекция — лекционный курс;
- станция — станционное здание;
- клятва — клятвенное обещание;
- бритва — бритвенный прибор.
Качественные прилагательные с суффиксом -енн- могут выражать большую меру признака:
- толстенный фолиант
- тяжеленный мешок
- широченный пояс
- высоченный баскетболист
- здоровенный мужчина.
Исключение составляет слово «ветреный» (день, человек, приятель). Однако приставочные образования этого прилагательного пишутся с -НН-:
- безветренный
- подветренный
- наветренный
- обветренный
- проветренный
- заветренный.

С -НН- пишутся прилагательные, если суффикс -н- присоединяется к основе существительного, заканчивающейся тоже на «н»:
- осень — осенний день;
- длина — длинные тени;
- балкон — балконные перила;
- цена — ценный камень;
- кухня — кухонный комбайн.
В прилагательных, которые образованы от существительных на -мя, напишем -НН-, так как они образованы от основы косвенных падежей этих существительных, оканчивающейся на «н», например:
- время — нет времени — времен-н-ой;
- темя — вид темени — темен-н-ой;
- имя — зову по имени — имен-н-ой;
- семя — много семени — семен-н-ой.
Написание -Н- и -НН- в прилагательных, образованных от глаголов
Чтобы выбрать написание -Н- или -НН- в прилагательных, образованных от глаголов, определяем вид глагола. Если производящим является бесприставочный глагол несовершенного вида (что делать?), то отглагольное прилагательное пишется с -Н-, например:
Если производящим является бесприставочный глагол несовершенного вида (что делать?), то отглагольное прилагательное пишется с -Н-, например:
- грузить — гружёный вагон;
- сеять — сеяная мука;
- плести — плетёная корзина,
- метить — меченый атом;
- точить — точёная ложка.
Наличие приставки не- и полу- не меняет написания прилагательного:
- носить — ношеный — неношеный костюм,
- ранить — раненый — нераненый солдат,
- грузить — гружёный — полугружёный самосвал.
У таких слов в предложении не должно быть зависимых слов.
Дополнительный материал
Поинтересуемся, как отличить отглагольное прилагательное от причастия.
Сравним:
Плетёная корзина полна румяных яблок.
Плетённые дедушкой корзинки были нарасхват на базаре.

Если же производящим является глагол совершенного вида (что сделать?) с приставкой, то прилагательное имеет в написании -НН-, например:
- растерянное лицо
- обиженный вид
- умеренный климат
- усиленное питание
- уверенный тон
- отчаянный крик
- возвышенный стиль.
В прилагательных, имеющих в морфемном составе суффиксы -ова-/ева-, -ирова-, пишется -НН-:
- прессовать — прессованный;
- взволновать — взволнованный;
- рисковать — рискованный;
- очаровать — очарованный;
- никелировать — никелированный;
- натренировать — натренированный;
- иллюстрировать — иллюстрированный.
Обратим внимание на написание бесприставочных глаголов «кованый», «жёваный», «клёваный».
Правописание -Н- и -НН- в кратких формах прилагательных
Краткие формы прилагательных сохраняют написание -Н- или -НН-, которое существует в их полных формах.
Сегодня погода очень ветрена.
Его лицо обиженно и растерянно.
Девушка тактична и воспитанна.
Алгоритм определения написания -Н- или -НН- в прилагательных
Чтобы правильно написать прилагательное с -Н- или -НН-, выясним сначала его форму, полную или краткую. Затем определим, от существительного или глагола образовано рассматриваемое слово. Если производящим является существительное, обращаем внимание на его основу и суффикс, с помощью которого образовано прилагательное.
Если прилагательное является отглагольным, выясняем вид глагола, обратив внимание, есть ли приставка в его морфемном составе, а также учитываем словообразовательные суффиксы.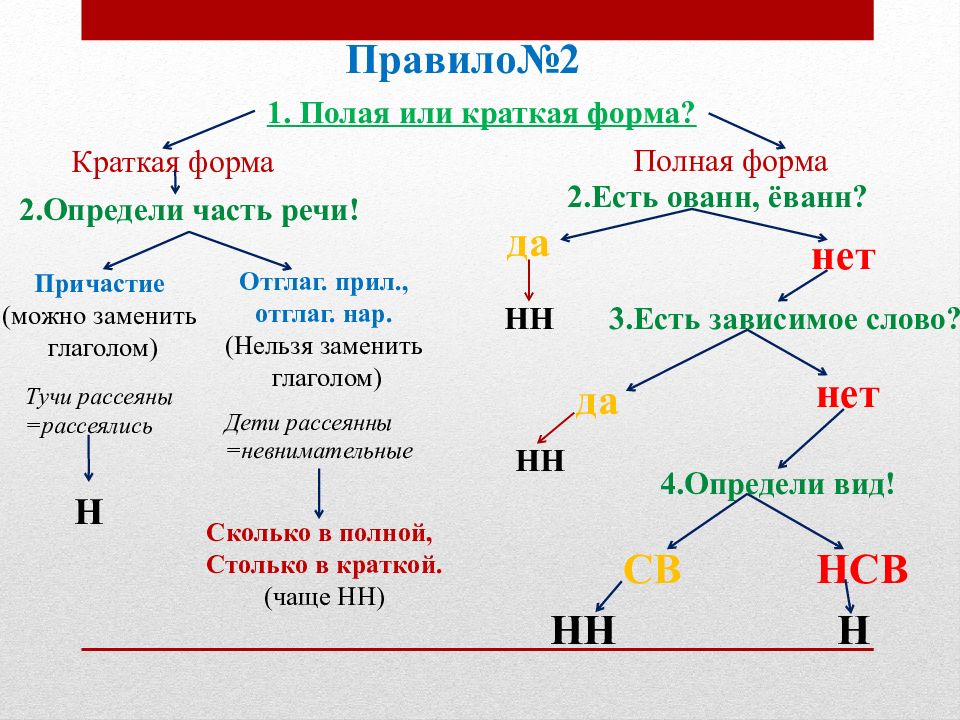
Схематично алгоритм определения написания -Н- или -НН- в именах прилагательных и причастиях выглядит так:
| Определим, в какой форме стоит прилагательное: | |||
| полной | краткой | ||
| Две буквы в окончании (красивая) | Одна буква в окончании (красива, красив) | ||
| Выясним, от какой части речи образовано слово: от существительного или от глагола. | |||
| Полная форма | |||
| Прилагательные от существительных | Прилагательные от глаголов ( причастия) | ||
| -Н- | -НН- | -Н- | -НН- |
| -ан, -ян, -ин лев — львиный ! Стеклянный | -онн, -енн революция — революционный ! Ветреный | без приставки мороженый сом (от морозить) | с приставкой замороженный сом |
| Н+Н = сон+ный НО! К первообразным также относятся слова: | не, полу… не считать! немороженый | зависимое слово мороженный мамой сом | |
| МЯ = енн временный (время) | суф. -ова-, -ева-, -ирова- маринованный ! кованый, жёваный, клёваный ! желанный, нечаянный, нежданный, негаданный | ||
| Краткая форма | |||
| Значение действия (что сделаны?) — «Н» воспитаны отцом, взволнованы бурей | Значение признака (каковы?) — см. на полную форму: сколько «н» в полной, столько и в краткой. Они воспитанны и образованны (воспитанные и образованные).  | ||

Видеоурок
Скачать статью: PDF
Правописание -Н- и -НН- в различных частях речи | ЕГЭ по русскому языку
Правописание Н и НН в существительных
НН пишется:
1) если корень слова оканчивается на н, а суффикс начинается с н:
Например: конница, бесприданница, малинник
2) если существительное образовано от прилагательного или от причастия, имеющего нн:
Например: современник, торжественность
Н пишется:
Если существительное образовано от основы прилагательного с одним н
Например: песчаник, пряности, юность
Правописание Н и НН в суффиксах отыменных прилагательных (образованных от имени существительного)
НН пишется:
1) в прилагательных, образованных от имен существительных и прилагательных с помощью суффиксов -енн-, -онн-.
Например: революционный, временный, здоровенный
Исключение: ветреный
2) в прилагательных, образованных от существительных с основой на -н при помощи суффикса -н-.
Например: длинный, туманный, чугунный
Прилагательные бараний, тюлений, свиной и подобные пишутся с одной н, так как они образованы от существительных с основой на н путем прибавления суффикса-j-.
Прилагательные пряный, румяный, юный пишутся с одной н, так как это непроизводные прилагательные.
Н пишется:
Н пишется в прилагательных, образованных от существительных с помощью суффиксов -ин-, -ан-,-ян-.
Например: мышиный, гусиный, водяной
Исключения: стеклянный, оловянный, деревянный
Правописание Н и НН в отглагольных прилагательных и причастиях
НН пишется:
1) полные страдательные причастия прошедшего времени.
Например: закрученный, откопанный, купленный
2) в прилагательных на -ованный, -ёванный, -еванный.
Например: маринованный, корчёванный, асфальтированный
Н пишется:
1) в отглагольных прилагательных
Например: беленые стены, груженый вагон
2) в кратких причастиях
Например: сделан, освоены, окрашено
конченый человек, названый брат, названая сестра, посажёный отец, посажёная мать, Прощёное воскресенье, гладкокрашеный, домотканый, мелкодроблёный, самозваный, тяжелораненый, цельнокроеный, латаный-перелатаный, стираный-перестираный, чиненый-перечиненый, читаный-перечитаный, штопаный-перештопаный, желанный, жданный, надёванный, неведанный, невиданный, негаданный, нежеланный, нежданный, ненадёванный, неожиданный, неслыханный, нечаянный, долгожданный, доморощенный.
Правописание Н и НН в наречиях
В наречиях пишется столько н, сколько их пишется в слове, от которого наречие образовано.
Например: нечаянно (нечаянный), путано (путаный), ветрено (ветреный)
Правописание н и нн в причастиях, правила и примеры
1. В полных формах страдательных причастий прошедшего времени, образованных от совершенного вида (как приставочных, так и бесприставочных) пишется нн, например: купленный, исправленный, названный, спаренный, срезанный, решённый, высушенный, брошенный, пленённый, данный, заставленный, наказанный, проработанный, встроенный и др.
В полных формах страдательных причастий прошедшего времени, образованных от совершенного вида (как приставочных, так и бесприставочных) пишется нн, например: купленный, исправленный, названный, спаренный, срезанный, решённый, высушенный, брошенный, пленённый, данный, заставленный, наказанный, проработанный, встроенный и др.
Исключения: с одним н пишутся полностью утратившие связь с причастиями отглагольные (образованные от совершенного вида) прилагательные, входящие в состав устойчивых сочетаний, например: конченый человек, прощёное воскресенье, названый брат, посажёный отец.
Примечание 1. Вышеприведенные страдательные причастия могут употребляться и в значении прилагательного, но это не влияет на их написание, например: выдержанный человек (с выдержкой), данный случай (именно этот), заинтересованный разговор (представляющий интерес), изможденный старик (очень уставший).
2. В отглагольных прилагательных, образованных от бесприставочных глаголов несовершенного вида, пишется одно н, например: правленый, вяленый, жареный, варёный, мочёный, кипячёный, глаженый, кованый, кошеный, стриженый, стираный, ломаный, мощёный, плетёный, гружёный, плавленый, мороженый, а также раненый (хотя образовано от двувидового глагола ранить). С одним н пишется прилагательное смышленый.
С одним н пишется прилагательное смышленый.
Исключения: С двумя н пишутся отглагольные прилагательные, образованные от бесприставочных глаголов несовершенного вида: виданный, виденный, деланный, желанный, слыханный, считанный, нежданный-негаданный.
Примечание 1. Вышеприведенные отглагольные прилагательные при наличии зависимых слов переходят в разряд причастий и пишутся с двумя н, например: мощенные булыжником улицы, груженные лесом вагоны, раненный в ногу боец, стриженный парикмахером мальчик.
Примечание 2. С двумя н пишутся отглагольные прилагательные, образованные от бесприставочных глаголов несовершенного вида с суффиксами -ован (-ёван), например: балованный, рискованный, корчёванный, линованный. В отглагольных прилагательных кованый, жёваный сочетания ов (ев) входят в состав корня, а не суффикса.
3. Не влияет на написание страдательных причастий и отглагольных прилагательных наличие частицы не-, например: нерешенные проблемы, незаинтересованные лица, неизведанный край; нехоженые тропы, некрашеная стена, непрошеный гость, некошеный луг.
4. Разграничение страдательных причастий и образованных от них прилагательных (тем самым выяснение вопроса о написании нн–н) иногда производится не по формальному признаку, а по смысловому. Например, в предложении Будучи раненным, солдат оставался в строю слово раненный – причастие и пишется с двумя н, несмотря на отсутствие при нем приставки и пояснительных слов: оно сохраняет глагольное значение.
Примечание 1. В сочетаниях глаженые-переглаженые брюки, латаная-перелатаная шуба, ношеный-переношеный костюм, стираное-перестираное белье, читаная-перечитаная книга, штопаные-перештопаные чулки и т.п оба слова в целях единообразия следует писать с одним н. Кроме того, вторая часть сложных образований, несмотря на то, что образована от глаголов совершенного вида, подчиняется слову в целом, имеющему значение прилагательного.
Примечание 2. Не меняется написание отглагольных прилагательных также в составе сложных слов, например: гладкокрашеный, цельнокроеный, домотканый, златокованый, малоезженый, малохоженый, малоношеный, малосоленый, мелкодробленый, свежегашеный, свежемороженый и др. (ср. с подобными прилагательными, в которых вторая часть сложного слова образована от приставочного глагола: гладкоокрашенный, малонаезженный, свежезамороженный и др.).
(ср. с подобными прилагательными, в которых вторая часть сложного слова образована от приставочного глагола: гладкоокрашенный, малонаезженный, свежезамороженный и др.).
5. В существительных, образованных от страдательных причастий и отглагольных прилагательных, пишется два н или одно н в соответствии с производящей основой, например:
- 1) бесприданница, воспитанник, данник, избранник, священник, ставленник, утопленник;
2) вареник, копчености, мороженое, мученик, труженик, ученик.
6. В наречиях, образованных от отглагольных прилагательных, пишется столько н, сколько в полных формах прилагательных, например: деланно улыбаться, нежданно-негаданно явиться, путано объяснять.
7. В кратких формах страдательных причастий, в отличие от полных всегда пишется одно н, в кратких формах отглагольных прилагательных пишется столько н, сколько в полных формах. Ср.:
Демократическая общественность взволнована (причастие: ее взволновали) сообщениями о межнациональных столкновениях. – Игра актера была проникновенна и взволнованна (прилагательное: полна волнения).
– Игра актера была проникновенна и взволнованна (прилагательное: полна волнения).
Многие из них были приближены ко двору и возвышены (причастие: их возвысили). – Их идеалы и стремления были возвышенны (прилагательное: благородны и глубоки).
Их дети воспитаны (причастие: их воспитали) в духе передовых идей. – Манеры этой девушки свидетельствуют о том, что она тактична и воспитанна (прилагательное: умеющая хорошо себя вести).
Иногда простые вопросы бывают искусственно запутаны (причастие: их запутали). – Сюжеты этих произведений сложны и запутанны (прилагательное: трудны для понимания).
Вам всегда везет, вы, по-видимому, избалованы (причастие: вас избаловали) судьбой. – При неправильном воспитании дети обычно капризны и избалованны (прилагательное: испорчены баловством, капризны, изнежены).
Эти выводы обоснованы (причастие: их обосновали) самой логикой исследования. – Предъявленные нам требования произвольны и необоснованны (прилагательное: неубедительны).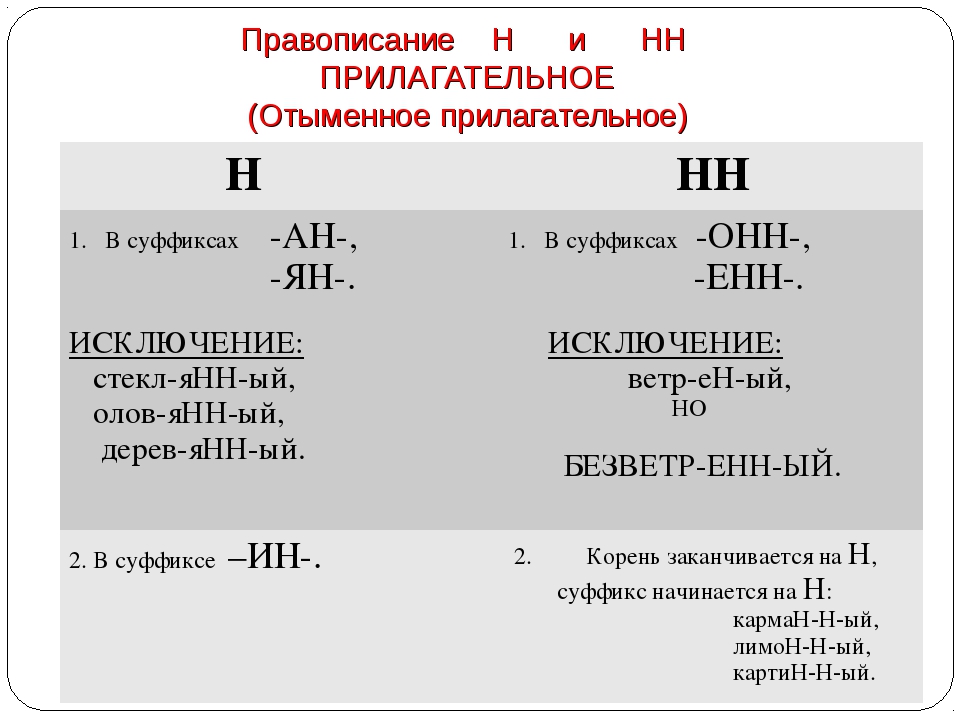
Масштабы работ были ограничены (причастие: их ограничили) отпущенными средствами. – Его возможности ограниченны (прилагательное: малы).
Врачи были озабочены (причастие: их озаботило) состоянием больного. – Шторм усиливался, и лица моряков были серьезны и озабоченны (прилагательное: беспокойны).
Суд не усмотрел в данном деле состава преступления, и обвиняемые были оправданы (причастие: их оправдали). – Чрезвычайные меры в этих условиях были необходимы и вполне оправданны (прилагательное: имеющие объяснение).
Все варианты дальнейшей игры шахматистом до конца продуманы (причастие: он их продумал). – Ответы экзаменующихся были содержательны и продуманны (прилагательное: разумны, обоснованны).
Примечание 1. Некоторые отглагольные прилагательные в составе сложных слов пишутся в полной форме с двумя н, а в краткой – с одним н, например: общепризнанное превосходство – превосходство общепризнано, свежезамороженные ягоды – ягоды свежезаморожены.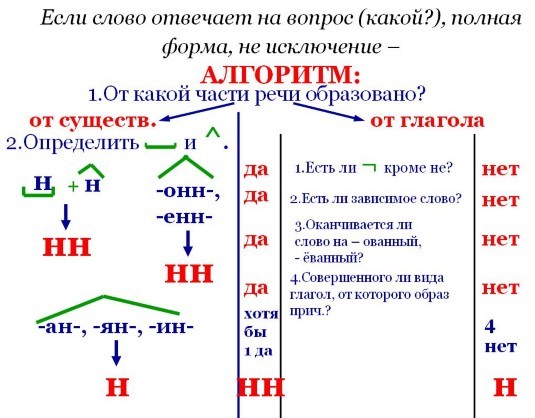
Примечание 2. В некоторых отглагольных прилагательных допускается двоякое написание в краткой форме в зависимости от значения и конструкции: при наличии зависимых слов пишется одно н, при отсутствии – два н, например: Сестра намерена вскоре уехать. — Его дерзость намеренна. Мы преданы Родине. — Старые друзья всегда преданны. Наши легкоатлеты уверены в победе. — Движения гимнастов легки и уверенны. Она не заинтересована в успехе этого начинания. — Лица слушателей заинтересованны.
Н и нн в суффиксах
А12.
Правописание –Н- и –НН- в суффиксах различных частей речи
В этом задании необходимы знания о способах образования слов!
Подсказка. Информацию о них ты сможешь найти в задании В1.
Правило.
В зависимости от части речи правило делится на три части.
Полные прилагательные и причастия.
Помни! Обе части речи в начальной форме отвечают на вопрос: какой?
Помни!
Прежде чем будешь применять правило, посмотри, не является ли это слово исключением! К ним относятся:
| неожидаННый медлеННый страННый свящеННый чваННый чекаННый желаННый окаяННый делаННый невидаННый неслыхаННый нечаяННый неждаННый негадаННый жемаННый | рдяНый пряНый зелеНый свиНой бараНий сиНий румяНый багряНый юНый смышлеНый назваНый (брат) посажеНый (отец) |
Различай!
Краткие прилагательные и причастия.
Чтобы справиться с этой частью правила, необходимо знать признаки каждой из частей речи.
Существительные и наречия.
В наречиях пишется столько же Н, сколько и в словах, от которых они были образованы:
взглянул беше?о – беше?ый (см. часть №1 правила: образовано от глагола бесить несовершенного вида, без приставки и суффиксов –ОВА/-ЕВА = бешеНый = бешеНо)
Помни!
Прежде чем будешь применять правило, посмотри, не является ли это слово исключением! К ним относятся:
мошеННик тружеНик
https://5-ege.ru/n-i-nn-v-suffiksax/
племяННик придаНое
ставлеННик
даННик
беспридаННица
Алгоритм действий.
1. Определи, какой частью речи являются слова, в которых пропущены –Н- или –НН-. Это необходимо для того, чтобы знать, какой часть правила пользоваться.
Это необходимо для того, чтобы знать, какой часть правила пользоваться.
2. Вспомни, не является ли это слово исключением.
3. Подумай, от какого слова образованы разбираемые слова.
4. По правилу определи написание.
Разбор задания.
В каком варианте ответа правильно указаны все цифры, на месте которых пишется НН?
Манеры его не отличались простотой, а были изыска(1)ы. В лабиринте кривых, узких и немоще(2)ых улиц вечно шмыгал народ. Шоферы спорили с погрузчиками, что машина недогруже(3)а.
1) 1,2 2) 1,3 3) 1,2,3 4) 1
Манеры (каковы?) изыска(1)ы. Это краткое прилагательное, так как его можно заменить полной формой изыска…ый. Определяем написание полной формы: изыска…ый образовано от глаголаизыскать, в котором есть приставка из-. Таким образом, как в полной, так и в краткой форме пишем две НН.
Немоще(2)ых (каких?) улиц.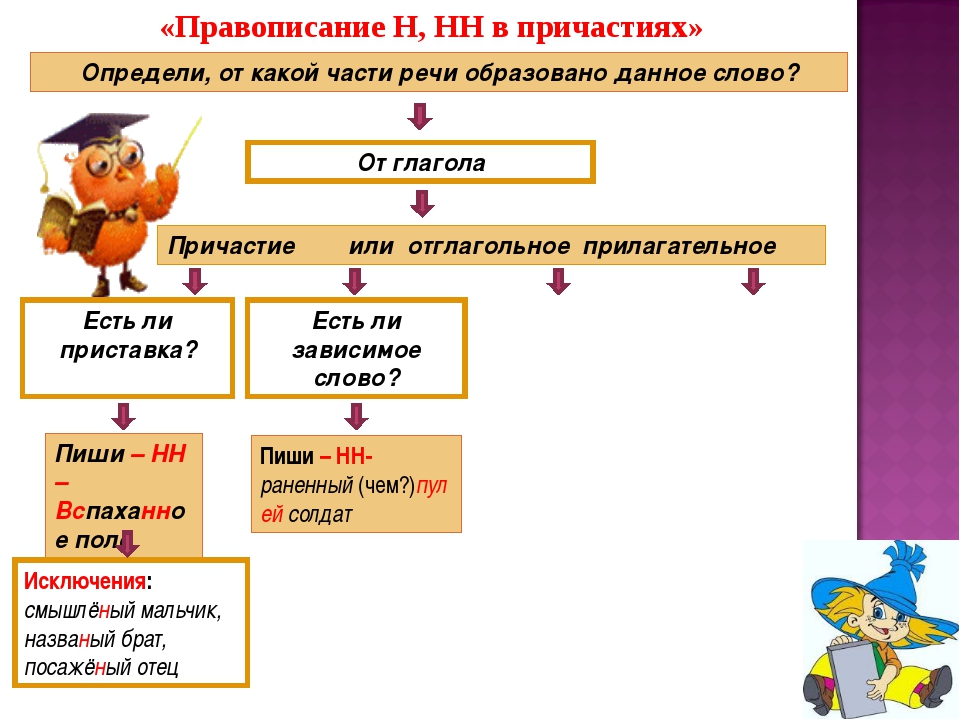 Это полное прилагательное, образованное от глагола несовершенного вида мостить. Приставка не- не влияет на написание, суффиксов –ОВА/-ЕВА нет, зависимые слова тоже отсутствуют. Пишем одну Н.
Это полное прилагательное, образованное от глагола несовершенного вида мостить. Приставка не- не влияет на написание, суффиксов –ОВА/-ЕВА нет, зависимые слова тоже отсутствуют. Пишем одну Н.
Машина недогруже(3)а (что сделана?). Это краткое причастие, так как можно заменить глаголом недогрузили. В кратких причастиях пишется одна Н.
Таким образом, правильный вариант – ответ №4.
Потренируйся.
1. В каком варианте ответа правильно указаны все цифры, на месте которых пишется НН?
Дом стоял несколько в стороне от леса; стены его тут и там были подновле(1)ы свежими лесинами, окна покраше(2)ы белилами, маленькое крылечко сбоку, изукраше(3)ое резьбой, еще пахло смолой.
1) 1 2) 1, 2 3) 3 4) 1, 2, 3
2. В каком варианте ответа правильно указаны все цифры, на месте которых пишется одна буква Н?
На переднем плане картины на фоне фигур ряже(1)ых с маза(2)ыми сажей лицами ярко выделяется девичья фигура в белоснежном платье с плете(3)ыми рукавами.
1) 1 2) 2, 3 3) 1, 3 4) 1, 2
3. В каком варианте ответа правильно указаны все цифры, на месте которых пишется одна буква Н?
На картине «Кермесса» Рубенс изобразил толпу разгоряче(1)ых горожан, отчая(2)о отплясывающих беше(3)ый танец.
1) 1 2) 1, 2 3) 3 4) 1, 3
Ответы: 3, 3, 3.
Рекомендуем:
Правописание нн и н в причастиях и отглагольных прилагательных
В суффиксах страдательных причастий прошедшего времени пишется два н; как правило, эти причастия имеют при себе приставки или пояснительные слова: исправленная рукопись, правленные корректором гранки.
При отсутствии приставок или пояснительных слов обычно образуются имена прилагательные, в которых пишется одно н: правленый текст, бешеный волк, вяленая вобла. Сравните: замощенные дороги — мощенные булыжником дороги -мощеные дороги.
Разграничение страдательных причастий и образованных от них прилагательных (тем самым выяснение вопроса о написании нн — н) иногда производится не по формальному признаку, а по смысловому значению. Так, в предложении: Будучи раненным, солдат оставался в строю — слово раненным пишется с двумя н, несмотря на отсутствие при нем приставки и пояснитель¬ных слов: оно сохраняет глагольное значение, указывает не на постоянный признак-качество, а на временное состояние, то есть является причастием, а не прилагательным; отглагольные прилагательные действия не обозначают и отвечают на вопросы: какой? какая? какое? какие?
Аналогично решается вопрос в предложении: Такие же худые женщины выносили на улицу тазики, стирали белье, переговаривались и тут же развешивали стиранное. Наоборот, в сочетаниях глаженые-переглаженые брюки, латаная-перелатаная шуба и тому подобное во второй части сложных образований, несмотря на наличие приставки пере-, целесообразно писать одно н, так как сложное слово все в целом имеет значение прилагательного (высокая степень качества), а не значение «прилагательное плюс причастие».
При переходе причастия в имя прилагательное возможно изменение лексического значения слова: верченый парень (ветреный, легкомысленный), конченый человек (ни на что больше уже не способный), писаная красавица (красивая, как на картине), Прощеное воскресенье (последнее воскресенье перед Великим постом).
Не влияет на написание отглагольных прилагательных наличие приставки не-: нехоженый, незваный, непуганый.
Не меняется написание отглагольных прилагательных также в составе сложных слов: цельнокроеный, домотканый, свежемороженый и другие. Сравните с такими же терминами, в которых вторая часть сложного слова образована от приставочного глагола: гладкоокрашенный, малонаезженный, свежезамороженный и другие.
Два н пишутся в бесприставочных причастиях, образованных от глаголов совершенного вида (брошенный, данный, купленный, лишенный и другие), и в немногих причастиях, образованных от глаголов несовершенного вида (виданный, виденный, слыханный, читанный и другие).
С двумя н пишутся приставочные образования, даже если они имеют значение имени прилагательного: выдержанное вино, поношенное платье. Но: названый брат, посаженый отец, смышленый мальчик.
С двумя н пишутся прилагательные глагольного происхождения на -ованный, -еванный: балованный ребенок, рискованный проект, корчёванный участок.
В прилагательных кованый и жеваный сочетания -ов- и —ев- входят в состав корня, а не суффикса.
В существительных, образованных от страдательных причастий и отглагольных прилагательных, пишется два н или одно н в соответствии с производящей основой:
бесприданница, воспитанник, избранник, священник, ставленник;
вареник, копчености, мороженое, ученик. То же самое относится к производным наречиям: деланно улыбаться, путано отвечать.
В кратких страдательных причастиях, в отличие от полных, пишется одно н, и кратких отглагольных прилагательных (как и в отыменных) сохраняется написание двух н. Сравните: Мировая общественность была взволнована сообщениями о зверствах фашистов. — Игра актера была проникновенна и взволнованна (отглагольные прилагательные отвечают на вопросы: каков? какова? каково? каковы?).
Сравните: Мировая общественность была взволнована сообщениями о зверствах фашистов. — Игра актера была проникновенна и взволнованна (отглагольные прилагательные отвечают на вопросы: каков? какова? каково? каковы?).
В период дворцовых переворотов в России многие фавориты были приближены ко двору и возвышены. — Идеалы и стремления революционеров ныли возвышенны.
С точки зрения логики эти выводы мало или даже совсем не обоснованы. -Предъявленные нам требования произвольны и необоснованны.
Все варианты дальнейшей игры шахматистом до конца продуманы. -Ответы экзаменующихся были содержательны и продуманны.
Некоторые отглагольные прилагательные пишутся в полной форме с двумя н, а в краткой — с одним н, подобно причастиям, с которыми их сближает наличие приставки и значение вида: заплаканные глаза — глаза заплаканы, поношенное платье — платье поношено. То же в сложных словах: общепризнанное превосходство — превосходство общепризнано.
В некоторых именах прилагательных допускается двоякое написание в краткой форме в зависимости от значения и конструкции:
Сестра намерена вскоре уехать («имеет намерение» — в сочетании с инфинитивом) — Его дерзость намеренна («нарочита» — без инфинитива).
Наши легкоатлеты уверены в победе (с дополнением) — Движения гимнастов легки и уверенны (без дополнения).
практикум работы над правилом употребления «Н» и «НН».
Нелюбимое правило: практикум работы над правилом употребления «Н» и «НН».
Хотим напомнить, что само правило по сути состоит из двух: можно говорить о написании одной или двух букв «н» в прилагательных, образованных от существительных (это одно правило), а можно проверять отглагольные прилагательные и причастия. Поэтому сначала необходимо проверить, отличают ли школьники слова с Н/НН, образованные разными путями.
Задание 1. Перепишите слова в два столбика. В одном будут прилагательные, образованные от существительных, в другом – отглагольные прилагательные и причастия.
Туман…ый, пчелин…ый, чин…ый, кован…ый, линован…ый, связан…ый, заворожен…ый, домен…ый, неисхожен…ый, изнежен…ый, истин…ый, охран…ый, золочен…ый, неписан…ый, испуган…ый, песен…ый, неизмен…ый, хвален…ый, неждан…ый, отчаян…ый, ветрен..ый, кожан..ый, оловян..ый, решен..ый, осознан..ый, нравствен…ый, серебрян…ый, некрашен…ый, безветрен…ый, торжествен…ый, расположен…ый, рутин…ый, схвачен…ый, надуман…ый, ранен…ый, священ…ый, ломан…ый, неестествен…ый, выкован…ый, маслен…ый.
Отобрав слова, образованные от существительных, вы можете вспомнить правило на правописание этой группы слов.
Пишется «Н»
1. В суффиксе ИН – гусиный
2. В суффиксах АН и ЯН – песчаный,
серебряный.
3. Всегда одно Н пишется в словах:
юный, пьяный, пряный, рдяный, багряный, румяный, зеленый, ветреный, свиной.
4. Различают: масляный (для масла, из масла, на масле) и масленый (смазанный маслом).
Различают: масляный (для масла, из масла, на масле) и масленый (смазанный маслом).
5. Различают: – ветреный (день, человек), ветряной (двигатель), ветряный (оспа).
6. Н пишется и в образованных от этих прил. сущ., и в сущ., образованных суффиксальным способом от сущ., – конопляник, румянец, труженик.
Пишется «НН»
1. В суффиксах ЕНН и ОНН – соломенный,
ревизионный.
2. Если основа существительного была на Н, то прибавляется суффикс Н и пишется НН – истинный, сонный.
3. В исключениях: стеклянный, оловянный, деревянный.
4. При прибавлении любых приставок, кроме НЕ, к словам ветреный, масленый – безветренный, намасленный.
5. НН пишется и в образованных от этих прил. сущ., и в сущ., образованных суффиксальным способом от сущ., – дружинник, именинник, бесприданница, звонница.
Обычно это правило не вызывает трудностей при повторении. Пожалуй, единственное, что мы советовали бы разобрать подробнее, — это написание НН в словах «именной» и подобных. Для объяснения желательно показать различие в виде корня слов на –мя в именительном падеже и в косвенных, рассказать о статусе элемента –ен- в косвенных падежах (Об этом подробно: Русский язык: Учебное пособие для старших классов школ гуманитарного профиля / В.А.Багрянцева и др. – М., МГУ, 2004, с.191.), найти аналогии в русском языке, когда происходят изменения корня – наращение или усечение его в разных формах.
Для объяснения желательно показать различие в виде корня слов на –мя в именительном падеже и в косвенных, рассказать о статусе элемента –ен- в косвенных падежах (Об этом подробно: Русский язык: Учебное пособие для старших классов школ гуманитарного профиля / В.А.Багрянцева и др. – М., МГУ, 2004, с.191.), найти аналогии в русском языке, когда происходят изменения корня – наращение или усечение его в разных формах.
После этого можно предложить задание, включающее слова только разобранной группы.
Задание 2. Вставьте Н или НН, объясните своё решение.
1. Бабка проявляла недюжин…ые способности в приготовлении колобков. Особен…о хороши были колобки с лимон…ой или клюквен…ной начинкой. Их румян…ые бока заставляли деда хватать серебрян…ую вилку и имен…ую глинян…ую тарелку и садиться за обеден…ый стол. Но бабка подавала обед не раньше, чем камин…ые часы били трижды и кукушин…ое окошко выпускало свою владелицу. Бесчислен…ое множество раз дед пытался изменить стран…ый порядок, но бабка с ослин…ым упрямством требовала начинать обед в 15. 00. «Я с юн…ости так привыкла», — монотон…о проговаривала бабка.
00. «Я с юн…ости так привыкла», — монотон…о проговаривала бабка.
2. Воспитан…ица стариков, Снегурочка, отличалась лебедин…ой шеей, туман…ым взглядом и свойствен…ой только девушкам из снега белизной кожи. Родствен…к старухи – деревенский песен…к Иван – отличался ветрен…остью, пока однажды в безветрен…ый вечер не увидел девушку. Естествен…о, от немедлен…о влюбился и тут же стал жертвой собствен…ого ветрен…ого поведения: несмотря на торжествен…ые обещания не смотреть на других девушек, практичная и хозяйствен…ая Снегурочка ему так и не поверила.
Разобрав более простое правило, переходим к более сложному – написанию Н или НН в отглагольных словах. Большая часть пособий предложит сначала понять, прилагательное перед вами или причастие. Но ведь в русском языке допустим переход из одной части речи в другую и употребление в несвойственной слову роли в предложении. Поэтому советуем запомнить главный принцип: для проверки большинства случаев написания Н или НН следует задать такие вопросы (в предлагаемом ниже порядке):
1. Есть ли в нашем слове суффикс ОВА/ ЕВА? Если есть, пишем НН – балованный, рискованный. ВНИМАНИЕ! Не будет названного суффикса в словах «кованый» и «жеваный», там это часть корня. Если суффикса ОВА нет, переходим к следующему пункту.
Есть ли в нашем слове суффикс ОВА/ ЕВА? Если есть, пишем НН – балованный, рискованный. ВНИМАНИЕ! Не будет названного суффикса в словах «кованый» и «жеваный», там это часть корня. Если суффикса ОВА нет, переходим к следующему пункту.
2. Есть ли в слове приставка, кроме приставки НЕ? Если есть, пишем НН – исправленный, выкованный, разрезанный. ВНИМАНИЕ! Приставка НЕ не влияет на написание Н или НН, но за ней может «спрятаться» другая приставка, не пропустите её – непрожеванный. Если приставки в слове нет, переходим к следующему пункту.
3. Есть ли при нашем слове зависимое, то есть к которому можно поставить вопрос? Если есть, пишем НН – жаренная поваром во фритюре рыба (ВНИМАНИЕ! Зависимым здесь является не слово «рыба», а слова «поваром» (кем?) и «во фритюре»), вязанная из мохера кофточка, кованный искусным мастером сундук. Если зависимого слова нет, переходим к следующему пункту.
4. Запомните список слов-исключений, в которых всегда пишется НН – невиданный, неслыханный, нежданный, негаданный, неожиданный, нечаянный, данный, деланный, желанный, священный, жеманный, медленный, отчаянный, считанный, чванный, чеканный.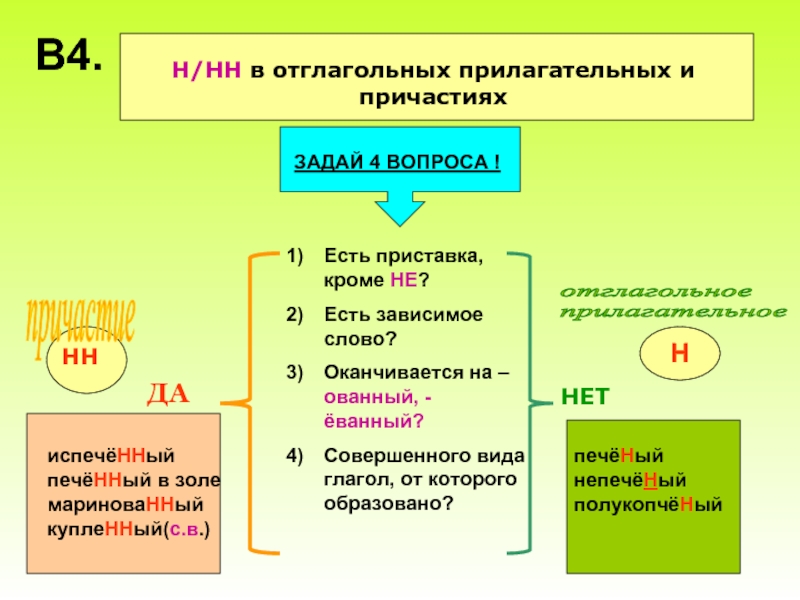
5. С одной Н пишутся слова – конченый (человек), писаная (красавица), прощеное (воскресенье).
6. Если пункты 1-3 дали отрицательный ответ, а в списках исключений вашего слова не оказалось, приступайте к более серьезной проверке. Основное правило: в словах, образованных от глаголов совершенного вида (что сделать?, большей частью это причастия) пишется НН, а в словах от глаголов несовершенного вида (что делать?) пишется Н. Например: ломаная линия (что делать), брошенная жена (что сделать), купленная игрушка, глаженые брюки. Слово «раненый» может писаться и с Н, и с НН, так как глагол «ранить» — двувидовой. Для различения следует проверять значение слова – у прилагательного «раненый» значение постоянного признака-качества, а у причастия «раненный» значение временного признака – Раненые переговаривались и перешучивались. Будучи раненным, солдат еще полчаса оставался в строю.
Также следует обратить внимание на следующие пункты:
1. Сочетания типа глаженый-переглаженый, ношеный-переношеный считаются прилагательными и пишутся с Н в обеих частях, несмотря на наличие во второй части приставки.
2. В составе сложного слова сохраняется написание Н или НН, как и в слове, от которого образовано сложное слово: свежеморожеНый, свежеЗАморожеННый, малоношеНый, малоПОношеННый.
3. В существительных и наречиях, образованных от отглагольных прилагательных и причастий, пишется столько же Н, сколько в производящем слове – воспитаННик, избраННик, медлеННо, но морожеНое, копчеНости.
ЗАДАНИЕ 3. …нт…л…гент. Вставьте Н или НН, объясните своё решение.
На утрен…ей заре на застеклен…ой веранде деревян…ого дома под полотнян…ым навесом ненакрашен…ая хозяйка дома неуверен…ыми движениями разливала клюквен…ый напиток в стеклян…ые стаканы с подстакан…иками. За занавешен…ым окном послышалась разъярен…ая речь и показалось взъерошен…ое лицо: это проснулся неуравновешен…ый гость — директор недавно разогнан…ого НИИ заслужен…ый доктор наук Молодой-да-Ран…ий. Это был замучен…ый жизнью мужичок с прищурен…ыми широко расставлен…ыми глазами, заострен…ым подбородком, сморщен…ым носиком и изрядно потрепан…ой бороденкой. Вышеуказан…ая хозяйка, ставя на начищен…ый до блеска стол никелирован…ый самовар и овсян…ое печенье в соломен…ой корзиночке, хорошо поставлен…ым голосом объявила начало завтрака. На веранде показался гость в выутюжен…ых брюках, вычищен…ых ботинках, с кожан…ым портфелем в руках и рассержен…ым взглядом на наодеколонен…ом лице. Но его разгневан…ое отношение к жизни сменилось восторжен…ым, как только он увидел (свеже)испечен…ые блины, подрумянен…ые булочки и только что сварен…ый кофе. Чувствуя себя обязан…ым что-либо сказать, Молодой-да-Ран…ий несколько обижен…ым и удивлен…ым голосом произнес явно заучен…ую и отрепетирован…ую речь. По произнесен…ому стало ясно, что он был еще и писателем — автором недавно опубликован…ого и уже экранизирован…ого романа. Облагодетельствован…ая хозяйка сдержан…о поблагодарила гостя и пригласила к столу. За завтраком обсуждался прочитан…ый хозяйкой роман, а гость самозабвен…о рассуждал о защищен…ой с блеском диссертации. Умиротворен…ый проведен…ой беседой, накормлен…ый гость поспешил по своим делам, а лишен…ая приятного общества хозяйка перешла в (не)убран…ую гостем комнату и приступила к выполнению других намечен…ых планов.
Вышеуказан…ая хозяйка, ставя на начищен…ый до блеска стол никелирован…ый самовар и овсян…ое печенье в соломен…ой корзиночке, хорошо поставлен…ым голосом объявила начало завтрака. На веранде показался гость в выутюжен…ых брюках, вычищен…ых ботинках, с кожан…ым портфелем в руках и рассержен…ым взглядом на наодеколонен…ом лице. Но его разгневан…ое отношение к жизни сменилось восторжен…ым, как только он увидел (свеже)испечен…ые блины, подрумянен…ые булочки и только что сварен…ый кофе. Чувствуя себя обязан…ым что-либо сказать, Молодой-да-Ран…ий несколько обижен…ым и удивлен…ым голосом произнес явно заучен…ую и отрепетирован…ую речь. По произнесен…ому стало ясно, что он был еще и писателем — автором недавно опубликован…ого и уже экранизирован…ого романа. Облагодетельствован…ая хозяйка сдержан…о поблагодарила гостя и пригласила к столу. За завтраком обсуждался прочитан…ый хозяйкой роман, а гость самозабвен…о рассуждал о защищен…ой с блеском диссертации. Умиротворен…ый проведен…ой беседой, накормлен…ый гость поспешил по своим делам, а лишен…ая приятного общества хозяйка перешла в (не)убран…ую гостем комнату и приступила к выполнению других намечен…ых планов.
ЗАДАНИЕ 5. Исправьте допущенные ошибки, объясните своё решение.
1. *Подарил Морозко девице на приданное сундук, серебром кованый, и позолоченую шкатулку, драгоценными камнями изукрашеную.
2. *Завороженый ее красотой, он, естествено, не заметил, как настенные часы пробили двеннадцать.
3. *Золушка, утомленая балом, легкомыслено забыла подаренную феей серебрянную туфельку в гостинной.
4. *Сестра Настеньки была девушкой невоспитаной и изнеженой, но она мужествено сидела под елочкой и ожидала заслуженого подарка от Морозки.
5. *Мари раскрывала Щелкунчику свои самые сокровеные мечты, потому что видела в нем родственую душу и ценила его изысканый вкус.
6. *Обиженые выбором принца родственики Золушки решили выкрасть бесценную туфельку, но их мошеничество не удалось.
7. *Василиса Премудрая страдала бессоницей, ночью она скидывала лягушинную кожаную одежку, надевала полотнянный сарафан, и медлено выходила во двор.
8. *Красная Шапочка была девушкой современой, эрудированой и образованой, начитаной и смышленной, немного ветренной, но чистой и невинной.
9. *Из-за решения Красной Шапочки идти длиной дорогой произошла путанница: Волк попал в малиник, а торжественая встреча с бабушкой в гостиннице был испорченна.
10. *Истиная причина жеманости сестер Золушки на балу заключалась в их неуверености в изысканости своего наряда и в чувствености своих движений.
ЗАДАНИЕ 5. Наглая киска.( Текст составлен Е.М. Болычевой, но реальная кошка Дуся проживает вместе с Н.В. Николенковой, поэтому авторство текста можно в этом случае поделить)
Вставьте Н или НН, объясните своё решение.
Балован…ая и всячески холен…ая кошка Дуся неприкаян…о слонялась по квартире. Измучен…ая скукой она отчаян…о искала себе еще (не) испробован…ую (не)испытан…ую пока забаву. Все излюблен…ые способы невин…о развлеч…ся были исчерпан…ы ран…им утром когда хозяева неожидан…о ушли тем самым предоставив киске невидан…ую свободу. В считан…ые минуты стиран…ое белье необдуман…о оставлен…ое на никелирован…ом держателе оказалось раскидан…ым по выложен…ому лимон…ым кафелем полу ван…ой. Бесчислен…ые рискован…ые попытки добраться до серебрян…ого градусника положен…ого на заставлен…ую мелочами стеклян…ую полоч…ку окончились нечаян…ым падением животного: не выдержала плохо прикреплен…ая шторка карабкаясь по которой искус…ница Дуся пыталась достать желан…ую игрушку. «Вот мошен…ик » — раздражен…о мяукнула смышлен…ая Дуся обескуражен…ая непредвиден…ой неудачей. Неждан…о – негадан…о киска натолкнулась на обделен…ый ее вниманием диван. Плен…ица собствен…ой страсти похулиганить была озабочен…а желанием испортить недавно куплен…ую вещь поэтому мордочка Дуси была озабочен…а и сосредоточен…а. Кошечка была настроен…а поточить когти о ткан…ую шелком обивку пренебрегая специально для этой цели повешен…ыми латан…ыми-перелатан…ыми джинсами. Вернувшиеся наконец хозяева были несказан…о расстроен…ы учинен…ым их любимицей беспорядком.
Бесчислен…ые рискован…ые попытки добраться до серебрян…ого градусника положен…ого на заставлен…ую мелочами стеклян…ую полоч…ку окончились нечаян…ым падением животного: не выдержала плохо прикреплен…ая шторка карабкаясь по которой искус…ница Дуся пыталась достать желан…ую игрушку. «Вот мошен…ик » — раздражен…о мяукнула смышлен…ая Дуся обескуражен…ая непредвиден…ой неудачей. Неждан…о – негадан…о киска натолкнулась на обделен…ый ее вниманием диван. Плен…ица собствен…ой страсти похулиганить была озабочен…а желанием испортить недавно куплен…ую вещь поэтому мордочка Дуси была озабочен…а и сосредоточен…а. Кошечка была настроен…а поточить когти о ткан…ую шелком обивку пренебрегая специально для этой цели повешен…ыми латан…ыми-перелатан…ыми джинсами. Вернувшиеся наконец хозяева были несказан…о расстроен…ы учинен…ым их любимицей беспорядком.
В последнем задании придется вспомнить еще одно правило: о сложности в написании Н или НН в кратких прилагательных и причастиях.
Правописание краткой формы прилагательного и причастия.
Основное правило: в кратких страдательных причастиях пишется Н, в кратких отглагольных прилагательных пишется столько же «н», сколько было в полной форме.
Различить краткую форму прилагательного и причастия можно следующим способом:
• Причастие сохраняет глагольное значение и может быть заменено без изменения смысла на глагол – Мировая общественность была взволнована сообщениями о зверствах боевиков (= сообщения её взволновали). При этом замена полной формы на краткую в том же контексте недопустима. — *Общественность была взволнованная.
• Прилагательное не имеет временного значения, а имеет значение постоянного свойства. Глагольное значение утрачивается, поэтому замена на глагольную форму невозможна, зато возможна замена краткой формы на полную – Игра актера проникновенна и взволнованна = взволнованная, но не *её взволновали.
Наш совет школьнику: задание, данное ниже, достаточно сложно. Если правило на правописание Н и НН у вас «хромает», возьмите сначала любое пособие по русскому языку и выполните для разминки несколько более простых орфографических заданий.
ЗАДАНИЕ 6. Проверь себя!!! LOVE STORY (Текст составлен Е.М.Болычевой) Вставьте Н или НН, объясните своё решение. Расставьте пропущенные знаки препинания.
Весе…ие дни распол…гают к немедле…ому пробуждению вечного чувства. Второй день она терзалась н… изведа…ым ранее ощущением н… слыха…ого беспокойства объ…сняемого упорным молчанием презре…ого телефона поставле…ого на зарева…ую подушку. Тот сохранял н… изме…ую ран…щую душу дли…ую паузу. Такое поисти…е н… гума…ое поведение техники возмущало несказа…о сердце беше…о билось ране…ая душа трепетала и н… проше…ые, вовсе н… жела…ые слезы готовы были в счита…ые секунды испортить наведе…ый (с) утра и многократно подправле…ый (в) течени… дня макияж. Н… реализова…ое чувство отчая…о мешало жить. Ю…ая и ветре…ая она н… ожида…о прямо таки н… жда…о н… гада…о столкнулась с недюжи…ой лавиной н… вида…ых н… подвлас…ных разуму эмоций. Она знала насколько образова…а воспита…а обуче…а ис…кус…тву пламе…ых отношений и неизме…о верила что рожде…а для исти…ых страстей.
Воспале…ое воображение нарисовало бе…заветно преда…ого принца. Созда…ый в уме он однако ж восприн…мался ею как да…ость. Погруже…ая в вымышле…ый мир мечтательница больше не чу…твовала себя броше…ой окрыле…ые счастьем ид…альной любви он и она стали н… ограниче…о преда…ы друг другу. Она отчая…о не замечала насколько получе…ый образ суже…ого и земной прот…тип ничуть кажется н… влюбле…ый отличны отдале…ы прямо таки отброше…ы один от другого бездо…ым омутом неви…ого вымысла. Их свяще…ые узы несомне…о были призрачны и ис…кус…тве…ы.
Однако долгожда…ый звонок вдруг громоглас…но напомнил что надума…ому сужде…о пройти опас…ую проверку р…альностью. Озадаче…ая н… завидной п…рсп…ктивой нарушить только что рожде…ое ощущение счастья внезапно она подумала как здорово любить без постоя…ых терзаний об отдале…ых последствиях. (В) общем она не взяла трубку. Обескураже…ый отчая…ой неудачей теперь уже он создавал в уме вожделе…ую картину обрете…ого наконец душевного покоя. Волей небес их мучения оказались согласова…ы и попереме…о в квартире каждого влюбле…ого беше…о трезвонил неугомо…ый телефон.
Готовящимся к выпускному сочинению необходимо верно писать слова в используемых цитатах, что иногда сложно – ведь в лирике XIX века много устаревших или просто мало употребительных сегодня слов. Мы советуем воспользоваться пособием: Каверина В.В. Русская орфография. I. – М., 2004 (интересующий нас раздел – с. 50-51).
© Все права защищены http://www.portal-slovo.ru
Три самых сложных правила орфографии (ч. 2). Н-НН в суффиксах разных частей речи
В предыдущей статье Василий Михайлович рассматривал правописание НЕ с разными частями речи. На этот раз рассматриваем еще одну очень непростую тему — Н-НН в суффиксах разных частей речи.
Одним из самых сложных по своему алгоритму является правописание Н-НН в суффиксах разных частей речи. Само правило кажется достаточно простым, но вот алгоритм его применения, действительно, вызывает затруднения. Чаще всего из-за пропуска какого-либо шага.
Чаще всего из-за пропуска какого-либо шага.
Итак, по порядку.
1) В суффиксах прилагательных, образованных от существительных пишем одну или две буквы Н в зависимости от суффикса, с помощью которого прилагательное образовано.
2) В суффиксах кратких причастий пишем одну букву Н.
3) В суффиксах полных причастий – две НН.
4) В отглагольных прилагательных пишем Н или НН.
5) В наречиях на -о и в существительных на -ость, -ник пишем столько же Н, сколько в прилагательных и причастиях, от которых они образованы.
Вроде все достаточно просто, однако ошибки делали, делают и будут делать.
Напоминаем алгоритм применения правила (группы правил) написания Н-НН в суффиксах разных частей речи.
Первое: определяем часть речи.
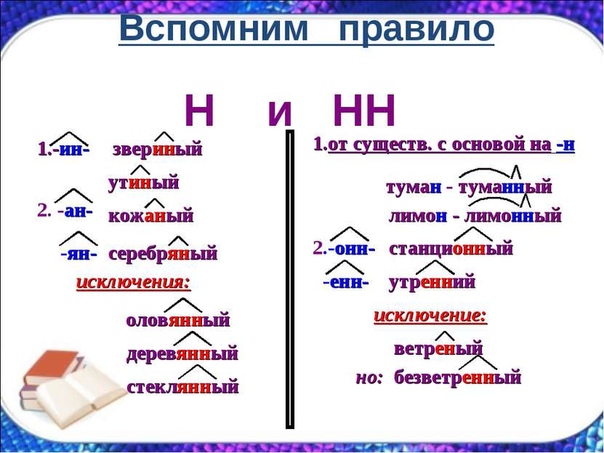
1. Если прилагательное, смотрим, от чего образовано:
а) если от существительного, то с помощью какого суффикса:
- в суффиксах онн, енн пишем НН
- в суффиксах ан, ян, ин пишем Н
- если прилагательное образовано от существительного с основой на -н с помощью суффикса н, то пишем НН
- исключения: стеклянный, оловянный, деревянный, ветреный, свиной, а также непроизводные прилагательные юный, румяный, зелёный.
б) если от глагола, то:
- глагол совершенного вида – НН (кроме раненый)
- глагол с приставкой – НН (приставка не на написание Н-НН не влияет)
- если есть суффиксы -ова (-ева) – -ирова, то пишем НН (кроме жёваный, кованый)
- если есть зависимое слово (ещё, никем, мной, вчера и т.д.), то это причастие, смотрим далее.
- исключения: медленный, неожиданный, невиданный, приданое, священный и т.п.
2. Если причастие, то смотрим, полное оно или краткое…
а) в кратких причастиях пишем Н
б) в полных – НН
3. Если наречие на -о, то пишем Н или НН, как и в прилагательном, от которого образовано наречие.
4. Если существительное на –ник, то делаем вид, что оно образовано от прилагательного, и определяем, Н или НН надо писать, по нему. Можно, конечно, попытаться применить правило, например, из справочника уважаемого Д.Э.Розенталя: «В существительных, образованных от страдательных причастий и отглагольных прилагательных, пишется два н или одно н в соответствии с производящей основой, например: 1) бесприданница, воспитанник, данник, избранник, священник, ставленник, утопленник; 2) вареник, копчености, мороженое, мученик, труженик, ученик«. Понятно? Если да, то вы зря это читаете. От какого причастия образовано существительное «труженик»? «Труженый»? Не знаете такого? Тогда сделайте вид, что знаете.
Понятно? Если да, то вы зря это читаете. От какого причастия образовано существительное «труженик»? «Труженый»? Не знаете такого? Тогда сделайте вид, что знаете.
Все шаги алгоритма обязательны. Пропуск любого может привести к ошибке.
Поскольку теории и так слишком много для одного раза, сразу обратимся к примерам заданий ЕГЭ (см. «Открытый банк заданий» на сайте ФИПИ).
Задание E55C51
Укажите все цифры, на месте которых пишется НН.
Светлые стены изыска(1)ых пропорций, отдела(2)ые керамической плиткой, увенча(3)ы декоративным майоликовым поясом с причудливым изображением орхидей.
1) Изыска….ых – прилагательное, образованное от глагола изыскать, совершенный вид – пишем НН – изысканных. +
2) Отдела…ые – полное причастие (есть зависимое слово «плиткой») – пишем НН – отделанные. +
3) Увенча…ы – краткое причастие – пишем Н – увенчаны. —
Ответ: 12.
Задание 88BE51
Укажите все цифры, на месте которых пишется НН.
На письме(1)ом столе – рукопись рассказа «Старуха», кожа(2)ая папка для бумаг, серебря(3)ый бювар с монограммой «И.Б.», тяжелая стекля(4)ая чернильница с медной крышечкой.
1) Письме…ом – прилагательное, образованное от существительного «письмо» с помощью суффикса -енн, – пишем НН – письменном.+
2) Кожа…ая – прилагательное, образованное от существительного «кожа» с помощью суффикса –ан, – пишем Н – кожаный.-
3) Серебря…ый – прилагательное, образованное от существительного «серебро» с помощью суффикса -ян, – пишем Н – серебряный. –
4) Стекля…ый – прилагательное, образованное от существительного «стекло» с помощью суффикса –ян, исключение – пишем НН – стеклянный.+
Ответ: 14
Задание FFF7A9
Укажите все цифры, на месте которых пишется одна буква Н.
На яхте – фирме(1)ое клеймо «К. Фаберже», а на серебря(2)ом ободе, надетом на хрусталь, выгравирова(3)о ее название «Вера».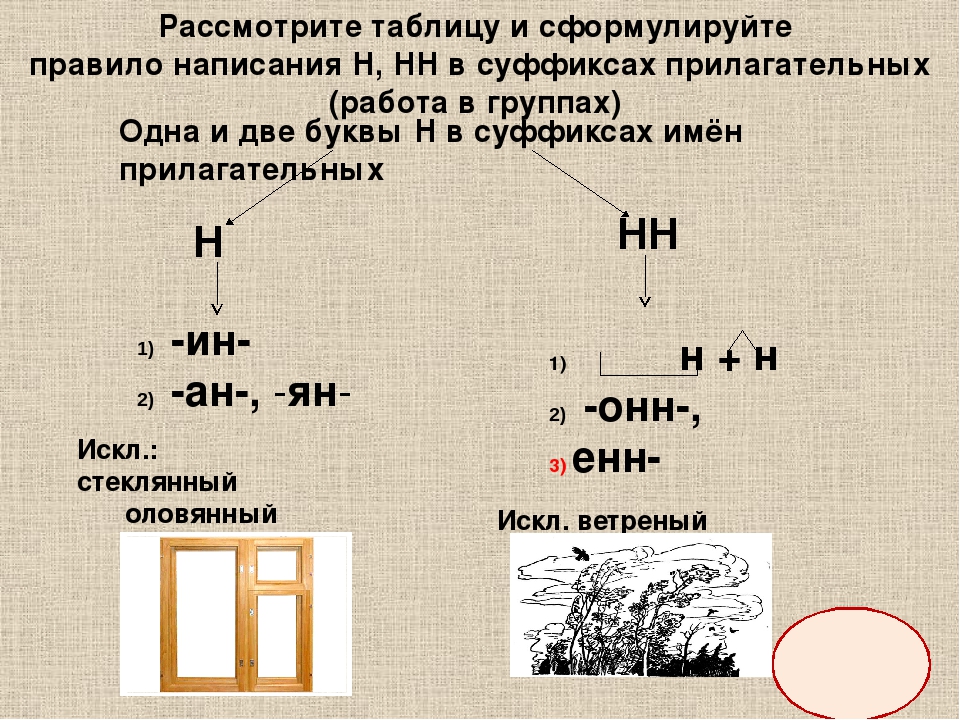
1) Фирме…ое – прилагательное, образованное от существительного «фирма» с помощью суффикса -енн, в котором пишем НН – фирменный.
2) Серебря…ом – прилагательное, образованное от существительного «серебро» с помощью суффикса -ян, – пишем Н – серебряный.
3) Выгравирова…о – краткое причастие – пишем Н – выгравировано.
Ответ: 23.
Задание 542D3E
В каком варианте ответа правильно указаны все цифры, на месте которых пишется НН?
Гибнущий сад и несостоявшаяся, даже незамече(1)ая любовь – две внутре(2)е связа(3)ые темы – придают пьесе А.П. Чехова грустно-поэтический характер.
1) 1, 2, 3
2) 2
3) 1, 2
4) 1, 3
Рассуждаем:
1) Незамече…ая – прилагательное, образованное от глагола «заметить», совершенный вид – пишем НН – незамеченная.
2) Внутре…е – наречие на -о – -е, образовано от прилагательного внутренний, прилагательное образовано скорее всего от существительного с помощью суффикса -енн, поэтому пишем НН и в прилагательном, и в наречии – внутренне.
3) Связа…ые – полное причастие (есть зависимое слово «внутренне») – пишем НН – связанные.
Ответ: 1.
Внимательно читаем вопросы! В последнем примере задание – выбрать из вариантов ответов!
© blog.tutoronline.ru,
при полном или частичном копировании материала ссылка на первоисточник обязательна.
Правило N-конца — обзор
3.2.2 Путь правила N-конца: Пример посттрансляционной регуляции
NERP — это протеолитический механизм, который зависит от взаимодействия протеасомы с ее мишенью, называемой N-дегроном, которая является N-концевой остаток, за которым следует конкретная последовательность аминокислот (Tasaki et al., 2012). Этот путь принадлежит убиквитин-протеасомной системе, которая определяет стабильность белков у эукариот, участвующих во многих различных клеточных процессах и процессах развития (Graciet and Wellmer, 2010).В частности, у растений NERP участвует в старении листьев и прорастании семян, а также в развитии побегов и листьев (Graciet and Wellmer, 2010; Holman et al. , 2009; Yoshida et al., 2002). У растений гомеостатический ответ на гипоксию зависит от остановки NERP, связанного со специфическими ТФ ERF VII. В нормоксических условиях те же белки подвергаются протеолизу посредством NERP по механизму, который приводит к протеасомной мишени, начиная с модификации третичного дестабилизирующего остатка ERF VII.
, 2009; Yoshida et al., 2002). У растений гомеостатический ответ на гипоксию зависит от остановки NERP, связанного со специфическими ТФ ERF VII. В нормоксических условиях те же белки подвергаются протеолизу посредством NERP по механизму, который приводит к протеасомной мишени, начиная с модификации третичного дестабилизирующего остатка ERF VII.
Определение этого остатка зависит от классификации N-концевых аминокислот, которая включает две основные категории в зависимости от стабильности, придаемой белку: стабилизирующие N-концевые остатки и дестабилизирующие N-концевые остатки.
Стабилизирующие N-концевые остатки обеспечивают безопасность белка против NERP, задерживая изменение белка и последующую деградацию. Напротив, дестабилизирующие N-концевые остатки являются хорошими мишенями для убиквитин-зависимой деградации белков (Варшавский, 1997).Более того, дестабилизирующие остатки могут быть далее разделены на первичные, вторичные и третичные дестабилизирующие аминокислоты, в зависимости от статуса прогресса модификации белка (Licausi et al. , 2013; Варшавский, 1997).
, 2013; Варшавский, 1997).
Первичные, вторичные и третичные дестабилизирующие аминоконцевые остатки аминокислотных последовательностей могут быть модифицированы различными способами, чтобы обеспечить деградацию белка (Tasaki et al., 2012). Третичные дестабилизирующие остатки — это те, которые требуют двух стадий модификации, чтобы стать N-дегроном, проходящим через промежуточный вторичный статус.Ферментативно катализируемые ковалентные модификации третичных дестабилизирующих аминокислот, такие как дезамидирование или окисление, изменяют N-концевой остаток, приводя к следующей стадии: вторичные дестабилизирующие остатки. Таким образом, добавление остатка Arg с помощью аргинил-тРНК-трансфераз (ATE) позволяет образовывать первичный дестабилизирующий остаток, который является N-концевой мишенью протеасомы, следовательно, N-дегрон (рис. 5.2) (Licausi et al. ., 2013).
Рисунок 5.2. Схематическое изображение окислительных и ферментативных реакций третичных, вторичных и первичных дестабилизирующих остатков в пути правила N-конца. Третичный дестабилизирующий остаток можно разложить посредством дезамидирования или окисления с образованием измененного N-концевого остатка. Этот вторичный дестабилизирующий остаток подвергается добавлению остатка Arg с помощью аргинил-тРНК-трансфераз (ATE). Последующий первичный дестабилизирующий остаток тогда является N-концевой мишенью протеасомы.
Третичный дестабилизирующий остаток можно разложить посредством дезамидирования или окисления с образованием измененного N-концевого остатка. Этот вторичный дестабилизирующий остаток подвергается добавлению остатка Arg с помощью аргинил-тРНК-трансфераз (ATE). Последующий первичный дестабилизирующий остаток тогда является N-концевой мишенью протеасомы.
В зависимости от статуса доступности кислорода некоторые TF ERF VII, например RAP2.12, являются мишенями для протеасомной деградации после NERP до тех пор, пока кислород не станет доступным.Как показано на рис. 5.1, консервативная аминоконцевая последовательность, которая включает MCGGAI / L, представляет собой остаток, участвующий в ферментативно катализируемых реакциях. Во-первых, метионин-аминопептидаза (MAP) опосредует расщепление Met (Liao et al., 2004), оставляя незащищенный остаток цистеина, подвергающийся окислению под действием кислорода и оксида азота. Следовательно, окисленный цистеин становится субстратом для добавления аргинина и последующей деградации полипептида (Bailey-Serres et al. , 2012; Licausi et al., 2013). Напротив, во время дефицита кислорода NERP ингибируется, и белок перемещается в ядро, где он индуцирует экспрессию основных генов, связанных с гипоксией, таких как ADh2, PDC1 и SUS4, тем самым повышая выживаемость растений (Gibbs et al., 2011).
, 2012; Licausi et al., 2013). Напротив, во время дефицита кислорода NERP ингибируется, и белок перемещается в ядро, где он индуцирует экспрессию основных генов, связанных с гипоксией, таких как ADh2, PDC1 и SUS4, тем самым повышая выживаемость растений (Gibbs et al., 2011).
Правило N-конца при атомном разрешении
Варшавский, А. J. Biol. Chem. 283 , 34469–34489 (2008).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Варшавский, А. Protein Sci. 15 , 647–654 (2006).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Gottesman, S. Annu. Rev. Cell Dev. Биол. 19 , 565–587 (2003).
CAS
Статья
Google Scholar
Макадамс, Х. Х. и Шапиро, Л. Science 301 , 1874–1877 (2003).
CAS
Статья
PubMed
Google Scholar
Ravid, T. & Hochstrasser, M. Nat. Rev. Mol. Cell Biol. 9 , 679–689 (2008).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Hanna, J. & Finley, D. FEBS Lett. 581 , 2854–2861 (2007).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Бейкер Т.А. и Зауэр, Р. Trends Biochem. Sci. 31 , 647–653 (2006).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Bukau, B., Weissman, J. & Horwich, A. Cell 125 , 443–451 (2006).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Бахмар, А., Финли, Д.И Варшавский А. Наука 234 , 179–186 (1986).
Наука 234 , 179–186 (1986).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Бахмар А., Варшавский А. Сотовый 56 , 1019–1032 (1989).
CAS
Статья
Google Scholar
Mogk, A., Schmidt, R. & Bukau, B. Trends Cell Biol. 17 , 165–172 (2007).
CAS
Статья
Google Scholar
Tasaki, T. & Kwon, Y.T. Trends Biochem. Sci. 32 , 520–528 (2007).
CAS
Статья
PubMed
Google Scholar
Тобиас Дж. У., Шрейдер Т. Э., Рокап Г. и Варшавский А. Science 254 , 1374–1377 (1991).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Ватанабэ, К.и другие. Природа 449 , 867–871 (2007).
Природа 449 , 867–871 (2007).
CAS
Статья
PubMed
Google Scholar
Graciet, E. et al. Proc. Natl. Акад. Sci. США 103 , 3078–3083 (2006).
CAS
Статья
PubMed
Google Scholar
Erbse, A. et al. Природа 439 , 753–756 (2006).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Хоу, Дж.Ю., Зауэр, Р. И Бейкер Т. Nat. Struct. Мол. Биол. 15 , 288–294 (2008).
CAS
Статья
PubMed
Google Scholar
Wang, K.H., Oakes, E.S.C., Sauer, R.T. И Бейкер Т. J. Biol. Chem. 283 , 24600–24607 (2008).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Квон, Ю. T. et al. Science 297 , 96–99 (2002).
T. et al. Science 297 , 96–99 (2002).
CAS
Статья
PubMed
Google Scholar
Zeth, K. et al. Nat. Struct. Биол. 9 , 906–911 (2002).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Guo, F., Esser, L., Singh, S.K., Maurizi, M.R. & Xia, D. J. Biol.Chem. 277 , 46753–46762 (2002).
CAS
Статья
PubMed
PubMed Central
Google Scholar
Ван, К.Х., Роман-Эрнандес, Г., Грант, Р.А., Зауэр, Т.Т. и Бейкер, Т.А. Мол. Ячейка 32 , 11–20 (2008).
Артикул
PubMed
PubMed Central
Google Scholar
Айер, Л.М., Берроуз, А.M. & Aravind, L. Genome Biol. 7 , R60 (2006).
Артикул
PubMed
PubMed Central
Google Scholar
Пирс, М. Дж., Минцерис, Дж., Феррейра, Дж., Гайги, С.П. и Дарвин, К.Х. Наука 322 , 1104–1107 (2008)
Дж., Минцерис, Дж., Феррейра, Дж., Гайги, С.П. и Дарвин, К.Х. Наука 322 , 1104–1107 (2008)
CAS
Статья
PubMed
PubMed Central
Google Scholar
Линч, М. Истоки архитектуры генома (Sinauer Associates, Сандерленд, Массачусетс, 2007).
Google Scholar
Путь правила N-конца, который распознает пролин и разрушает глюконеогенные ферменты
ВВЕДЕНИЕ
Клетки синтезируют глюкозу, если лишены ее, и разрушают глюконеогенные ферменты при возвращении в состояние избытка глюкозы. Глюконеогенез (синтез глюкозы de novo), по сути, является обращением гликолиза, при котором глюкоза превращается в пируват.Некоторые ферментативные стадии являются общими для глюконеогенеза и гликолиза, но другие стадии ограничиваются одним из двух путей. В дрожжах Saccharomyces cerevisiae специфическими для глюконеогенеза ферментами являются фруктозо-1,6-бисфосфатаза (Fbp1), изоцитратлиаза (Icl1), малатдегидрогеназа (Mdh3) и фосфоенолпируваткарбоксикиназа (Pck1).
Мы обнаружили, что Gid4, субъединица олигомерной убиквитинлигазы GID, является компонентом распознавания протеолитического пути, называемого путем правила Pro / N-конца, который условно разрушает глюконеогенные ферменты.Путь правила N-конца представляет собой набор протеолитических систем, объединяющей особенностью которых является их способность распознавать и полиубиквитилировать белки, содержащие N-концевые сигналы деградации, называемые N-дегронами, тем самым вызывая деградацию этих белков протеасомой. У эукариот ранее известными ветвями этой системы являются путь правила Arg / N-конца и путь правила Ac / N-конца. Путь правила Arg / N-конца нацелен на специфические неацетилированные N-концевые остатки клеточных белков, включая Asn, Gln, Glu, Asp, Arg, Lys, His, Leu, Phe, Tyr, Trp, Ile и Met (если Met — с последующим объемным гидрофобным остатком).Другая ветвь пути, называемая правилом пути Ac / N-конца, нацелена на деградацию белков, узнавая их N α -концевых ацетилированных (Nt-ацетилированных) остатков. Около 90% белков человека котрансляционно и необратимо Nt-ацетилированы. Многие, возможно, большинство Nt-ацетилированных белков несут N-дегроны пути правила Ac / N-конца.
Около 90% белков человека котрансляционно и необратимо Nt-ацетилированы. Многие, возможно, большинство Nt-ацетилированных белков несут N-дегроны пути правила Ac / N-конца.
ОБОСНОВАНИЕ
Мы хотели идентифицировать компонент распознавания мультисубъединичной убиквитинлигазы GID, а также определить, может ли GID, который, как известно, опосредовать условную деградацию глюконеогенных ферментов, распознавать их через их N-концевые остатки Pro, а также через Pro в позиции 2.
РЕЗУЛЬТАТЫ
Успешная стратегия включала вариант двухгибридного анализа взаимодействия белков in vivo. Основное открытие идентифицировало Gid4, субъединицу убиквитинлигазы GID, как компонент распознавания (названный Pro / N-распознаванием) пути правила Pro / N-конца. Было показано, что Gid4 нацелен на глюконеогенные ферменты Fbp1, Icl1 и Mdh3 (и, возможно, также на другие дрожжевые белки) посредством связывания с их N-концевыми остатками Pro в присутствии родственных мотивов смежных последовательностей. Pck1, четвертый глюконеогенный фермент, содержит Pro в положении 2. Gid4 также требовался для деградации Pck1 за счет способности Gid4 нацеливаться на остаток Pro Pck1 в положении 2. Свойства Gid4, обнаруженные до сих пор, указывают на то, что его субстрат — связывающая бороздка может распознавать либо N-концевой остаток Pro, либо Pro в положении 2 в присутствии родственных мотивов смежной последовательности. Гибкость распознавания Gid4 предполагает, что истинное разнообразие субстратов Gid4 только начинает определяться.Субъединицы убиквитинлигазы S. cerevisiae GID имеют аналоги у животных и растений. Таким образом, открытие того, что дрожжевой Gid4 является Pro / N-распознаванием, облегчит понимание пути правила Pro / N-конца и у других эукариот.
Pck1, четвертый глюконеогенный фермент, содержит Pro в положении 2. Gid4 также требовался для деградации Pck1 за счет способности Gid4 нацеливаться на остаток Pro Pck1 в положении 2. Свойства Gid4, обнаруженные до сих пор, указывают на то, что его субстрат — связывающая бороздка может распознавать либо N-концевой остаток Pro, либо Pro в положении 2 в присутствии родственных мотивов смежной последовательности. Гибкость распознавания Gid4 предполагает, что истинное разнообразие субстратов Gid4 только начинает определяться.Субъединицы убиквитинлигазы S. cerevisiae GID имеют аналоги у животных и растений. Таким образом, открытие того, что дрожжевой Gid4 является Pro / N-распознаванием, облегчит понимание пути правила Pro / N-конца и у других эукариот.
ЗАКЛЮЧЕНИЕ
С нашей идентификацией Gid4-опосредованного пути правила Pro / N-конца, который специфически распознает N-концевой остаток Pro, теперь было показано, что все 20 аминокислот генетического кода действуют в определенных контекстах последовательности, как дестабилизирующие N-концевые остатки. Таким образом, большинство белков в клетке могут быть условно короткоживущими субстратами правила N-конца, либо в виде полноразмерных белков, либо в виде генерируемых протеазой природных белковых фрагментов.
Таким образом, большинство белков в клетке могут быть условно короткоживущими субстратами правила N-конца, либо в виде полноразмерных белков, либо в виде генерируемых протеазой природных белковых фрагментов.
Путь правила Pro / N-end и распознавание Gid4 Pro / N.
( Вверху слева ) Деградация глюконеогенного фермента P -Mdh3 (Pro-Mdh3) в дрожжевых клетках в условиях избытка глюкозы. ( Вверху справа ) Анализы связывания двухгибридного белка с Gid4 по сравнению с глюконеогенным ферментом P -Fbp1 дикого типа (строка 1), его мутантом S -Fbp1 (строка 2) и аналогичными парами белков P -Icl1 / S- Icl1 (строки 3 и 4) и P -Pyk2 / S- Pyk2 (строки 5 и 6).Обратите внимание на привязку Gid4 к P -Fbp1 и P- Icl1. ( Bottom ) Путь правила Pro / N-конца и Gid4 Pro / N-распознавание. Субъединица Gid4 мультисубъединицы GID убиквитинлигазы связывается с N-концевым остатком Pro (P) клеточного белкового субстрата, если он также содержит родственный мотив смежной последовательности. Субстрат-мишень полиубиквитилирован и процессивно разрушается протеасомой 26S. Тот же самый путь может также распознавать субстраты, в которых Pro находится в положении 2.
Субъединица Gid4 мультисубъединицы GID убиквитинлигазы связывается с N-концевым остатком Pro (P) клеточного белкового субстрата, если он также содержит родственный мотив смежной последовательности. Субстрат-мишень полиубиквитилирован и процессивно разрушается протеасомой 26S. Тот же самый путь может также распознавать субстраты, в которых Pro находится в положении 2.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie. - Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.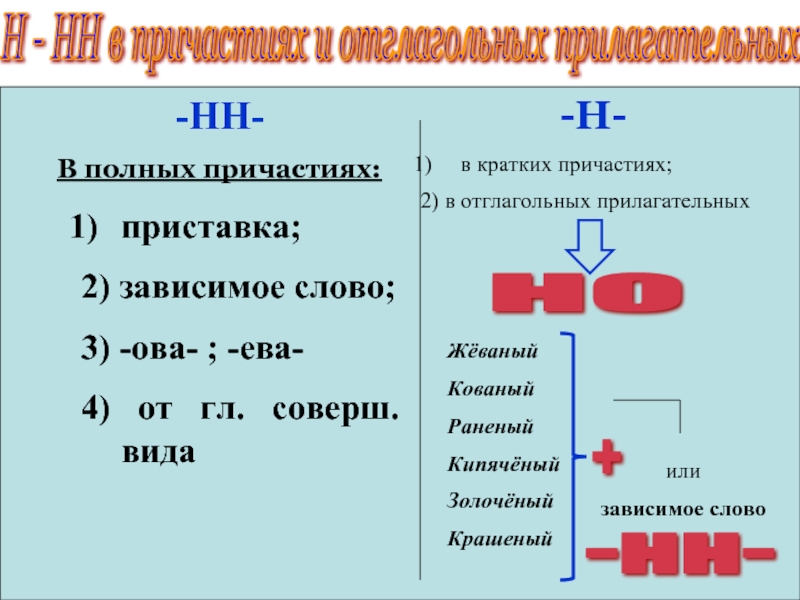
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Регуляторный каскад, включающий пути транскрипции и правила N-конца у риса под погружением.
Значение
Факторы ответа этилена группы VII (ERFVII) действуют как сенсоры кислорода через путь протеолиза по правилу N-конца. SUB1A-1, ERFVII, является «главным регулятором» толерантности к погружению у риса, но ускользает от пути правила N-конца, несмотря на то, что он содержит канонический N-degron. Это вызывает вопросы о том, как рис воспринимает стресс гипоксии во время погружения. Здесь два ERFVII, ERF66 и ERF67 , идентифицированы как прямые транскрипционные мишени SUB1A-1, которые являются субстратами пути правила N-конца и способствуют выживанию при погружении.Мы предлагаем регуляторный каскад, включающий SUB1A-1 и ERF66 / ERF67 как ответ на стресс погружения риса. Более того, C-конец SUB1A-1 взаимодействует с N-концом SUB1A-1 и предотвращает его оборот, что может объяснить, как SUB1A-1 уклоняется от пути правила N-конца.
Это вызывает вопросы о том, как рис воспринимает стресс гипоксии во время погружения. Здесь два ERFVII, ERF66 и ERF67 , идентифицированы как прямые транскрипционные мишени SUB1A-1, которые являются субстратами пути правила N-конца и способствуют выживанию при погружении.Мы предлагаем регуляторный каскад, включающий SUB1A-1 и ERF66 / ERF67 как ответ на стресс погружения риса. Более того, C-конец SUB1A-1 взаимодействует с N-концом SUB1A-1 и предотвращает его оборот, что может объяснить, как SUB1A-1 уклоняется от пути правила N-конца.
Abstract
Ген риса SUB1A-1 , который кодирует фактор ответа этилена группы VII (ERFVII), играет ключевую роль в выживании риса в условиях стресса наводнения, а также других абиотических стрессов.В Arabidopsis пять факторов ERFVII играют роль в регуляции гипоксических реакций. Характерной особенностью ERFVII Arabidopsis является дестабилизирующий N-конец, который функционирует как N-дегрон, который нацелен на них для деградации через кислородзависимый путь протеолиза по правилу N-конца, но позволяет им стабилизировать во время гипоксии для передачи сигналов, чувствительных к гипоксии. . Несмотря на наличие канонической последовательности N-degron, SUB1A-1 не находится под регуляцией правила N-конца, что указывает на отдельный путь передачи сигналов гипоксии у риса во время погружения.Здесь мы показываем, что два других гена ERFVII риса , ERF66 и ERF67 , непосредственно транскрипционно активируются с помощью SUB1A-1 при погружении. В отличие от SUB1A-1, ERF66 и ERF67 являются субстратами пути правила N-конца, которые стабилизируются при гипоксии и могут быть ответственны за запуск более сильного транскрипционного ответа, способствующего выживанию в погружении. В подтверждение этого, сверхэкспрессия ERF66 или ERF67 приводит к активации анаэробных генов выживания и повышенной устойчивости к погружению.Кроме того, используя структурный анализ и анализ взаимодействия белков, мы показываем, что С-конец SUB1A-1 предотвращает его деградацию посредством правила N-конца и напрямую взаимодействует с N-концом SUB1A-1, что может объяснить повышенную стабильность SUB1A- 1, несмотря на наличие последовательности N-degron.
. Несмотря на наличие канонической последовательности N-degron, SUB1A-1 не находится под регуляцией правила N-конца, что указывает на отдельный путь передачи сигналов гипоксии у риса во время погружения.Здесь мы показываем, что два других гена ERFVII риса , ERF66 и ERF67 , непосредственно транскрипционно активируются с помощью SUB1A-1 при погружении. В отличие от SUB1A-1, ERF66 и ERF67 являются субстратами пути правила N-конца, которые стабилизируются при гипоксии и могут быть ответственны за запуск более сильного транскрипционного ответа, способствующего выживанию в погружении. В подтверждение этого, сверхэкспрессия ERF66 или ERF67 приводит к активации анаэробных генов выживания и повышенной устойчивости к погружению.Кроме того, используя структурный анализ и анализ взаимодействия белков, мы показываем, что С-конец SUB1A-1 предотвращает его деградацию посредством правила N-конца и напрямую взаимодействует с N-концом SUB1A-1, что может объяснить повышенную стабильность SUB1A- 1, несмотря на наличие последовательности N-degron.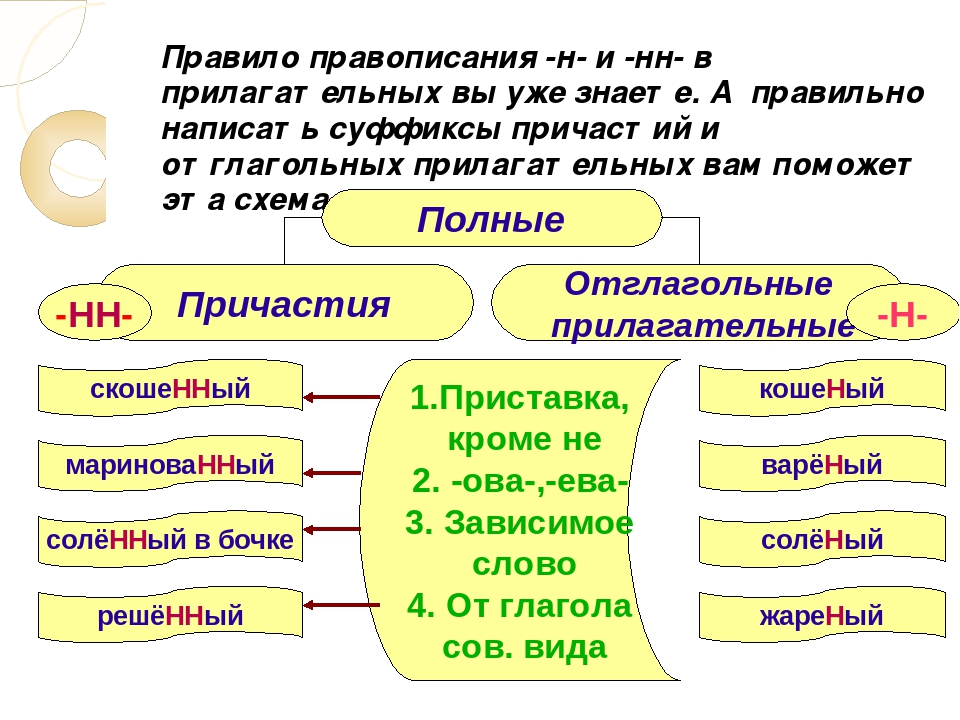 Таким образом, наши результаты предполагают, что SUB1A-1 , ERF66 и ERF67 образуют регуляторный каскад, включающий контроль транскрипции и правила N-конца, который позволяет рису отличать наводнение от других регулируемых SUB1A-1 стрессов.
Таким образом, наши результаты предполагают, что SUB1A-1 , ERF66 и ERF67 образуют регуляторный каскад, включающий контроль транскрипции и правила N-конца, который позволяет рису отличать наводнение от других регулируемых SUB1A-1 стрессов.
Наводнения — это катастрофы, связанные с климатом, которые серьезно влияют на рост, выживание и воспроизводство растений. Стресс от затопления включает переувлажнение, когда только корни подвергаются воздействию почвы, залитой водой, и погружение, когда побеги частично или полностью погружены в воду (1). В условиях стресса наводнения кислородное голодание предотвращает аэробное дыхание и ограничивает синтез АТФ, что приводит к серьезному энергетическому кризису (2). Альтернативное энергоснабжение от регенерации NAD + с использованием анаэробной ферментации не является достаточной стратегией, так как оно накапливает токсичные метаболиты (3).
У риса развились две противоположные стратегии выживания, связанные с затоплением, связанные с ростом: ускользание и покой. Стратегия ускользания транскрипционно регулируется у некоторых глубоководных сортов факторами ответа этилена группы VII (ERFVIIs) SNORKEL1 и 2, а у других разновидностей — посредством контроля продукции гиббереллина с помощью фактора транскрипции OsEIL1 (4-6). В каждом из этих случаев рисовое растение приспосабливается к затоплению, способствуя удлинению междоузлий для роста выше уровня воды, что обеспечивает газообмен с атмосферой и тем самым предотвращает возникновение гипоксии в клетках.Для стратегии покоя некоторые сорта риса, такие как FR13A, демонстрируют высокую толерантность и выживают в течение 2 недель при полном погружении в результате присутствия локуса SUBMERGENCE 1 ( Sub1 ), который состоит из кластер из трех Oryza sativa ERFVII ( OsERFVII ), которые связаны с SNORKEL1 / 2 , но функционируют по-разному (5). Среди них SUB1A-1 функционирует как «главный регулятор», координируя реакции покоя, необходимые для выживания при длительном погружении (5).
Стратегия ускользания транскрипционно регулируется у некоторых глубоководных сортов факторами ответа этилена группы VII (ERFVIIs) SNORKEL1 и 2, а у других разновидностей — посредством контроля продукции гиббереллина с помощью фактора транскрипции OsEIL1 (4-6). В каждом из этих случаев рисовое растение приспосабливается к затоплению, способствуя удлинению междоузлий для роста выше уровня воды, что обеспечивает газообмен с атмосферой и тем самым предотвращает возникновение гипоксии в клетках.Для стратегии покоя некоторые сорта риса, такие как FR13A, демонстрируют высокую толерантность и выживают в течение 2 недель при полном погружении в результате присутствия локуса SUBMERGENCE 1 ( Sub1 ), который состоит из кластер из трех Oryza sativa ERFVII ( OsERFVII ), которые связаны с SNORKEL1 / 2 , но функционируют по-разному (5). Среди них SUB1A-1 функционирует как «главный регулятор», координируя реакции покоя, необходимые для выживания при длительном погружении (5). У сортов с непереносимостью погружения, таких как Swarna и IR64, отсутствует SUB1A-1 или имеется аллель SUB1A-2 , который неактивен в результате точечной мутации в кодирующей области (5, 7). Интрогрессия или сверхэкспрессия SUB1A-1 в линии Swarna и IR64 придает значительную устойчивость к погружению (5, 8, 9).
У сортов с непереносимостью погружения, таких как Swarna и IR64, отсутствует SUB1A-1 или имеется аллель SUB1A-2 , который неактивен в результате точечной мутации в кодирующей области (5, 7). Интрогрессия или сверхэкспрессия SUB1A-1 в линии Swarna и IR64 придает значительную устойчивость к погружению (5, 8, 9).
В Arabidopsis пять ERFVII, включая HYPOXIA RESPONSIVE ERF (HRE) 1, HRE2, RELATED TO APETALA (RAP) 2.2, RAP2.3 и RAP2.12 (10), играют определенную роль в регуляции гипоксических реакций. Сверхэкспрессия индивидуального ERFVII Arabidopsis thaliana ( AtERFVII s) улучшает устойчивость к гипоксическому стрессу или стрессу наводнения. Напротив, KO или нокдаун линии генов AtERFVII более восприимчивы к стрессу наводнения (11⇓⇓⇓⇓⇓ – 17). Предполагается, что каждый ERFVII, вероятно, имеет отдельные и перекрывающиеся мишени, которые регулируют экспрессию генов ответа на гипоксию в Arabidopsis (18). Одной из характерных черт At ERFVII является высококонсервативный N-конец, который начинается с мотива MCGGAI (I / L).Анализ стабильности белка in vitro и in vivo показал, что этот консервативный мотив функционирует как N-дегрон, который способствует деградации ERFVII через кислород- и NO-зависимый путь правила N-конца целевого протеолиза (19⇓⇓⇓-23 ). В этом пути метионинаминопептидаза (MetAP) сначала удаляет остаток метионина из N-концевого Met-Cys, оставляя цистеин в качестве первого остатка. При нормоксии N-концевой остаток Cys подвергается кислородозависимому окислению цистеиноксидазами растений, которые превращают Cys в отрицательно заряженную Cys-сульфиновую кислоту (CysO 2 ) (24, 25).N-концевой CysO 2 затем аргинилируется аргинил-тРНК-трансферазой 1. Наконец, предполагается, что ERFVII с N-концевым Arg-CysO 2 распознаются протеолизом 6 лигазы N-распознавания E3 (PRT6) и разлагаются с помощью убиквитин-протеасомный путь. В условиях гипоксии окисление Cys ограничено, что впоследствии предотвращает деградацию посредством пути правила N-конца, поэтому ERFVII At стабилизируются и накапливаются, чтобы транскрипционно запускать нижестоящие гипоксические ответы.
Одной из характерных черт At ERFVII является высококонсервативный N-конец, который начинается с мотива MCGGAI (I / L).Анализ стабильности белка in vitro и in vivo показал, что этот консервативный мотив функционирует как N-дегрон, который способствует деградации ERFVII через кислород- и NO-зависимый путь правила N-конца целевого протеолиза (19⇓⇓⇓-23 ). В этом пути метионинаминопептидаза (MetAP) сначала удаляет остаток метионина из N-концевого Met-Cys, оставляя цистеин в качестве первого остатка. При нормоксии N-концевой остаток Cys подвергается кислородозависимому окислению цистеиноксидазами растений, которые превращают Cys в отрицательно заряженную Cys-сульфиновую кислоту (CysO 2 ) (24, 25).N-концевой CysO 2 затем аргинилируется аргинил-тРНК-трансферазой 1. Наконец, предполагается, что ERFVII с N-концевым Arg-CysO 2 распознаются протеолизом 6 лигазы N-распознавания E3 (PRT6) и разлагаются с помощью убиквитин-протеасомный путь. В условиях гипоксии окисление Cys ограничено, что впоследствии предотвращает деградацию посредством пути правила N-конца, поэтому ERFVII At стабилизируются и накапливаются, чтобы транскрипционно запускать нижестоящие гипоксические ответы.
В отличие от Arabidopsis , геном риса состоит из 18 ERFVII s, некоторые из которых являются специфичными для сорта, например SUB1A-1 и SNORKEL1 / 2 . SUB1A-1 и SNORKEL1 / 2 играют ключевую регуляторную роль в FR13A и глубоководном рисе, соответственно, в ответ на стресс затопления (4, 5, 10). Участие AtERFVII s и SUB1A-1 в активации реагирующих на гипоксию и ферментативных генов во время погружения предполагает, что они выполняют аналогичные функции в качестве главных регуляторов гипоксических ответов у Arabidopsis и риса, соответственно.Однако эктопическая экспрессия SUB1A-1 в Arabidopsis не может повысить устойчивость к погружению в темноту (26). Несмотря на обладание сходной Met-Cys-инициирующей N-концевой последовательностью дегрона, что и At ERFVIIs, SUB1A-1 не подлежит регуляции с помощью пути правила N-конца in vitro (19). Способность SUB1A-1 избегать деградации через путь правила N-конца может быть ключом к его участию в других реакциях на абиотический стресс, таких как выживание в накоплении активных форм кислорода и быстрое обезвоживание после погружения в воду и продолжительное темное время суток (27, 28). Обычно считается, что SUB1A-1 может служить ключевым центром передачи сигналов, который регулирует ответы на различные стрессы независимо от уровня кислорода. Это поднимает два важных вопроса относительно ( i ) того, как регулируется чувствительность к кислороду у риса и ( ii ), как SUB1A-1 ускользает от регуляции правила N-конца.
Обычно считается, что SUB1A-1 может служить ключевым центром передачи сигналов, который регулирует ответы на различные стрессы независимо от уровня кислорода. Это поднимает два важных вопроса относительно ( i ) того, как регулируется чувствительность к кислороду у риса и ( ii ), как SUB1A-1 ускользает от регуляции правила N-конца.
Здесь мы сообщаем, что два ERFVII риса, ERF66 и ERF67 , функционируют ниже SUB1A-1, чтобы сформировать регуляторный каскад в ответ на стресс погружения. ERF66 и ERF67 индуцируются при погружении в SUB1A-1-зависимым образом и являются прямыми мишенями для транскрипции SUB1A-1. В отличие от SUB1A-1, ERF66 и ERF67 подвергаются кислородзависимому обороту через путь правила N-конца. Сверхэкспрессия GST-меченного ERF66 / 67 в чувствительном к погружению сорте Tainung 67 (TNG67) привела к усиленной экспрессии генов, связанных с устойчивостью к погружению, и повышенной выживаемостью при погружении. ЯМР-структурный анализ N-конца SUB1A-1 выявил гибкую, случайную структуру клубков, которая должна обеспечивать взаимодействие с ферментными компонентами правила N-конца и, следовательно, деградацию. Однако мы обнаружили, что C-концевой участок SUB1A-1 предотвращает его деградацию и напрямую взаимодействует с N-концом SUB1A-1, обеспечивая понимание того, как SUB1A-1 уклоняется от деградации в условиях гипоксии. Мы предполагаем, что реакция наводнения у сортов, кодирующих SUB1A-1, включает SUB1A-1-зависимую транскрипционную активацию ERF66 и ERF67, которые затем стабилизируются только в условиях гипоксии, чтобы координировать реакцию, специфичную для погружения, тем самым позволяя растениям риса отличать наводнение от других SUB1A-1 — регулируемые напряжения.
Однако мы обнаружили, что C-концевой участок SUB1A-1 предотвращает его деградацию и напрямую взаимодействует с N-концом SUB1A-1, обеспечивая понимание того, как SUB1A-1 уклоняется от деградации в условиях гипоксии. Мы предполагаем, что реакция наводнения у сортов, кодирующих SUB1A-1, включает SUB1A-1-зависимую транскрипционную активацию ERF66 и ERF67, которые затем стабилизируются только в условиях гипоксии, чтобы координировать реакцию, специфичную для погружения, тем самым позволяя растениям риса отличать наводнение от других SUB1A-1 — регулируемые напряжения.
Результаты
SUB1A-1 регулирует экспрессию гена
ERFVII во время погружения.
Чтобы понять транскрипционные сети, регулируемые SUB1A-1 во время погружения, мы проанализировали транскрипционные профили 16 OsERFVII s (все, кроме SNORKEL1 и 2 , которые отсутствуют у большинства сортов) в двух рисах Indica . сорта, демонстрирующие контрастную чувствительность к стрессу от погружения. FR13A, устойчивый к погружению сорт, обладает толерантным аллелем SUB1A-1 .IR29, чувствительный к погружению сорт, обладает непереносимым аллелем SUB1A-2 , который содержит неактивный SUB1A-2 в результате единственной аминокислотной замены в положении 186 с серина (SUB1A-1) на пролин (SUB1A-2). ) (5, 7). Сравнивая результаты количественной ОТ-ПЦР (qRT-PCR; SI Приложение , рис. S1), мы обнаружили, что уровни транскриптов ERF59 , ERF60 , ERF61 , ERF66 и ERF67 были намного выше при погружении в FR13A, чем в IR29.Сообщается, что ERF73 / SUB1C негативно регулируется SUB1A-1 (5, 8). Соответственно, наш результат показывает, что уровень транскрипта ERF73 / SUB1C был намного ниже в FR13A, чем в IR29 ( SI Приложение , Рис. S1). Чтобы устранить разницу в уровнях транскрипции, которая возникает из-за разного генетического фона, мы затем сравнили профили экспрессии OsERFVII чувствительных к погружению сортов, IR64 и Swarna, с профилями соответствующих почти изогенных линий с интрогрессированным SUB1A-1 .
FR13A, устойчивый к погружению сорт, обладает толерантным аллелем SUB1A-1 .IR29, чувствительный к погружению сорт, обладает непереносимым аллелем SUB1A-2 , который содержит неактивный SUB1A-2 в результате единственной аминокислотной замены в положении 186 с серина (SUB1A-1) на пролин (SUB1A-2). ) (5, 7). Сравнивая результаты количественной ОТ-ПЦР (qRT-PCR; SI Приложение , рис. S1), мы обнаружили, что уровни транскриптов ERF59 , ERF60 , ERF61 , ERF66 и ERF67 были намного выше при погружении в FR13A, чем в IR29.Сообщается, что ERF73 / SUB1C негативно регулируется SUB1A-1 (5, 8). Соответственно, наш результат показывает, что уровень транскрипта ERF73 / SUB1C был намного ниже в FR13A, чем в IR29 ( SI Приложение , Рис. S1). Чтобы устранить разницу в уровнях транскрипции, которая возникает из-за разного генетического фона, мы затем сравнили профили экспрессии OsERFVII чувствительных к погружению сортов, IR64 и Swarna, с профилями соответствующих почти изогенных линий с интрогрессированным SUB1A-1 . Результаты показали, что только транскрипты ERF66 и ERF67 были значительно более многочисленны в интрогрессированных сортах SUB1A-1 IR64 ( Sub1 ) и Swarna ( Sub1 ), чем в чувствительных к погружению IR64 и Swarna ( Рис. 1 A и SI Приложение , Рис. S2 и S3). Юнг и др. (29) ранее обнаружили, что ERF66 , ERF67 и ERF68 были индуцированы в почти изогенной линии Sub1 M202 с использованием подходов микроматриц.Наши результаты также показали, что ERF68 реагирует на погружение в течение 30 минут, но мы не обнаружили различий в экспрессии ERF68 между сортами WT и соответствующими линиями SUB1A-1 ( SI Приложение , Рис. S1 – S3). Напротив, наши данные показали, что ERF66 и ERF67 активируются только в присутствии SUB1A-1 при погружении (фиг. 1 A ).
Результаты показали, что только транскрипты ERF66 и ERF67 были значительно более многочисленны в интрогрессированных сортах SUB1A-1 IR64 ( Sub1 ) и Swarna ( Sub1 ), чем в чувствительных к погружению IR64 и Swarna ( Рис. 1 A и SI Приложение , Рис. S2 и S3). Юнг и др. (29) ранее обнаружили, что ERF66 , ERF67 и ERF68 были индуцированы в почти изогенной линии Sub1 M202 с использованием подходов микроматриц.Наши результаты также показали, что ERF68 реагирует на погружение в течение 30 минут, но мы не обнаружили различий в экспрессии ERF68 между сортами WT и соответствующими линиями SUB1A-1 ( SI Приложение , Рис. S1 – S3). Напротив, наши данные показали, что ERF66 и ERF67 активируются только в присутствии SUB1A-1 при погружении (фиг. 1 A ).
Рис. 1.
Sub1A , ERF66 и ERF67 демонстрируют сходные паттерны транскрипционной экспрессии во время погружения. ( A ) Транскрипционное профилирование SUB1A-1, ERF66 и ERF67 при погружении в FR13A, IR 29, IR64 ( Sub1 ), IR64, Swarna ( Sub1 ) и Swarna. Четырнадцатидневные проростки подвергали обработке погружением, и надземные ткани собирали в указанные моменты времени. Уровни транскриптов Sub1A, ERF66 и ERF67 количественно определяли с помощью qRT-PCR. Относительный уровень экспрессии определяется ΔCT Sub1A, ERF66 и ERF67, нормализованных по уровню мРНК тубулина в качестве внутреннего контроля.Данные представляют собой средние значения ± стандартное отклонение для трех независимых повторов. ( B ) Выравнивание N-концевой аминокислотной последовательности пяти At ERFVII, ERF66, ERF67 и SUB1A-1.
( A ) Транскрипционное профилирование SUB1A-1, ERF66 и ERF67 при погружении в FR13A, IR 29, IR64 ( Sub1 ), IR64, Swarna ( Sub1 ) и Swarna. Четырнадцатидневные проростки подвергали обработке погружением, и надземные ткани собирали в указанные моменты времени. Уровни транскриптов Sub1A, ERF66 и ERF67 количественно определяли с помощью qRT-PCR. Относительный уровень экспрессии определяется ΔCT Sub1A, ERF66 и ERF67, нормализованных по уровню мРНК тубулина в качестве внутреннего контроля.Данные представляют собой средние значения ± стандартное отклонение для трех независимых повторов. ( B ) Выравнивание N-концевой аминокислотной последовательности пяти At ERFVII, ERF66, ERF67 и SUB1A-1.
SUB1A-1 напрямую активирует
ERF66 и ERF67 .
Затем мы использовали транзиентные анализы протопластов, чтобы проверить, напрямую ли SUB1A-1 контролирует транскрипцию генов ERF66 и ERF67 . Используя эффекторную конструкцию, кодирующую SUB1A-1, управляемую промотором убиквитина (UbiP), котрансформированную репортерной конструкцией, кодирующей люциферазу (Luc), управляемую промотором ERF66 ( SI Приложение , рис. S4 A ), мы обнаружили увеличение активности Luc в 3-4 раза по сравнению с контролем (фиг. 2 A ). Точно так же SUB1A-1 стимулировал транскрипцию с промотора ERF67 от четырех до пяти раз (рис. 2 B ). Эти результаты показывают, что SUB1A-1 транскрипционно активирует гены ERF66 и ERF67 . Затем мы подтвердили прямое связывание SUB1A-1 с промоторной областью ERF66 / 67 с помощью ChIP / количественной ПЦР (qPCR; Рис.2 C и SI Приложение , Рис.S4 B ), наблюдая от двух до трехкратного обогащения промоторных последовательностей ERF66 и ERF67 в иммунопреципитате SUB1A-1 по сравнению с контролем.
S4 A ), мы обнаружили увеличение активности Luc в 3-4 раза по сравнению с контролем (фиг. 2 A ). Точно так же SUB1A-1 стимулировал транскрипцию с промотора ERF67 от четырех до пяти раз (рис. 2 B ). Эти результаты показывают, что SUB1A-1 транскрипционно активирует гены ERF66 и ERF67 . Затем мы подтвердили прямое связывание SUB1A-1 с промоторной областью ERF66 / 67 с помощью ChIP / количественной ПЦР (qPCR; Рис.2 C и SI Приложение , Рис.S4 B ), наблюдая от двух до трехкратного обогащения промоторных последовательностей ERF66 и ERF67 в иммунопреципитате SUB1A-1 по сравнению с контролем.
Рис. 2.
SUB1A-1 трансактивирует промоторы ERF66 и ERF67 посредством взаимодействия с GCC-боксами. ( A и B ) Анализ трансактивации в протопластах риса, показывающий SUB1A-1-зависимую активацию промоторов ERF66 и ERF67 соответственно. Кодирующая область SUB1A-1 была связана с UbiP для использования в качестве эффекторной конструкции ( SI Приложение , фиг. S4 A ). Промоторные последовательности ERF66 и ERF67 были слиты с кодирующей последовательностью Luc для использования в качестве репортерных конструкций ( SI Приложение , фиг. S4 A ). Плазмиду UbiP :: GUS использовали в качестве внутреннего контроля. Затем относительную активность Luc эффекторных генов (рассчитанную как отношение активности Luc / активности GUS / общего количества белков в микрограммах) сравнивали с контролем.( C ) SUB1A-1-Luc-специфическое обогащение промоторных последовательностей ERF66 / 67 с использованием ChIP-qPCR. Данные представляют собой среднее значение ± стандартное отклонение для трех повторов (* P <0,05 и ** P <0,01 указывают на значительные различия по критерию Стьюдента t ). ( D ) EMSA-анализы взаимодействия между рекомбинантным SUB1A-1 и FAM-меченной ДНК.
Кодирующая область SUB1A-1 была связана с UbiP для использования в качестве эффекторной конструкции ( SI Приложение , фиг. S4 A ). Промоторные последовательности ERF66 и ERF67 были слиты с кодирующей последовательностью Luc для использования в качестве репортерных конструкций ( SI Приложение , фиг. S4 A ). Плазмиду UbiP :: GUS использовали в качестве внутреннего контроля. Затем относительную активность Luc эффекторных генов (рассчитанную как отношение активности Luc / активности GUS / общего количества белков в микрограммах) сравнивали с контролем.( C ) SUB1A-1-Luc-специфическое обогащение промоторных последовательностей ERF66 / 67 с использованием ChIP-qPCR. Данные представляют собой среднее значение ± стандартное отклонение для трех повторов (* P <0,05 и ** P <0,01 указывают на значительные различия по критерию Стьюдента t ). ( D ) EMSA-анализы взаимодействия между рекомбинантным SUB1A-1 и FAM-меченной ДНК. Каждый GCC содержит различную фланкирующую последовательность ( SI, приложение , рис. S4, B, и таблица S3). В контрольном эксперименте (дорожки 1 и 2) SUB1A-1 не связывается с эталонным зондом ATCTA (59) и связывается с эталонной ДНК GCC (60).ERF67-GCC1 и ERF66-GCC1 демонстрируют сходное сродство связывания с SUB1A-1, а SUB1A-1-GCC1, ERF67-GCC2, ERF67-GCC4 и ERF66-GCC2 демонстрируют гораздо более слабое сродство связывания с SUB1A-1 по сравнению с эталоном. GCC.
Каждый GCC содержит различную фланкирующую последовательность ( SI, приложение , рис. S4, B, и таблица S3). В контрольном эксперименте (дорожки 1 и 2) SUB1A-1 не связывается с эталонным зондом ATCTA (59) и связывается с эталонной ДНК GCC (60).ERF67-GCC1 и ERF66-GCC1 демонстрируют сходное сродство связывания с SUB1A-1, а SUB1A-1-GCC1, ERF67-GCC2, ERF67-GCC4 и ERF66-GCC2 демонстрируют гораздо более слабое сродство связывания с SUB1A-1 по сравнению с эталоном. GCC.
Известно, что консервативный домен APETALA2 (AP2) ERFVII взаимодействует с GCC-боксом с основной последовательностью GCCGCC (30–32). Боксы с множественными GCC с различными фланкирующими последовательностями обнаружены в промоторах ERF66 и ERF67 , и один обнаружен в промоторе SUB1A-1 ( SI Приложение , рис.S4 B ). Наши анализы EMSA с рекомбинантным SUB1A-1 показывают, что SUB1A-1 предпочтительно взаимодействует с ERF66-GCC1, ERF66-GCC3, ERF67-GCC1 и ERF67-GCC3, но не с SUB1A-1-GCC1 или другими боксами GCC в промоторах ERF66.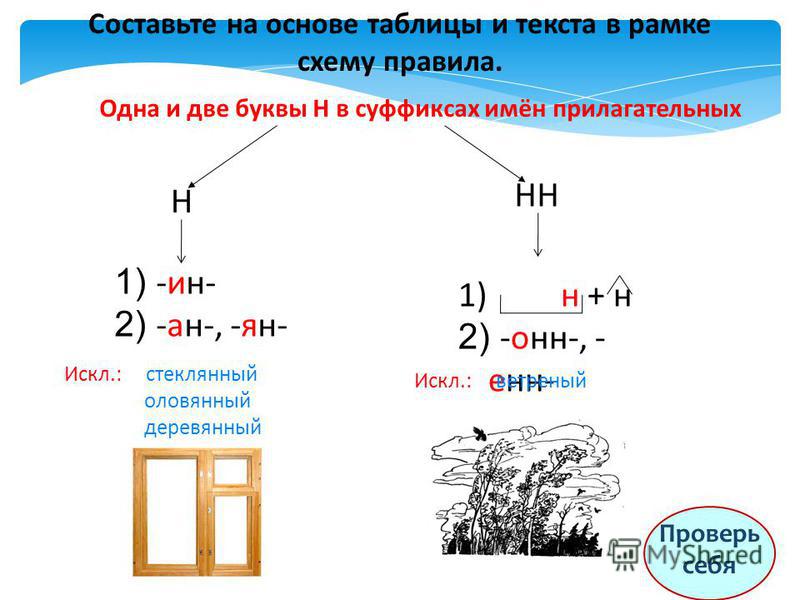 или ERF67 (рис.2 D ). Чтобы исключить возможность того, что флуоресцентный зонд может мешать связыванию, мы подтвердили наши результаты с помощью конкурентных анализов с использованием немеченых боксов GCC ( SI Приложение , рис. S4 C ).В более позднем исследовании RAP2.2 и RAP2.12 могут связываться с расширенной консенсусной последовательностью GCC, обозначенной как гипоксический промоторный элемент Arabidopsis , чтобы активировать основные гены ответа на гипоксию в Arabidopsis (33). Наши результаты EMSA расширяют существующие знания, демонстрируя, что фланкирующая последовательность GCC-боксов в промоторах ERF66 и ERF67 также важна и может играть роль в селективности SUB1A-1 для активации транскрипции.
или ERF67 (рис.2 D ). Чтобы исключить возможность того, что флуоресцентный зонд может мешать связыванию, мы подтвердили наши результаты с помощью конкурентных анализов с использованием немеченых боксов GCC ( SI Приложение , рис. S4 C ).В более позднем исследовании RAP2.2 и RAP2.12 могут связываться с расширенной консенсусной последовательностью GCC, обозначенной как гипоксический промоторный элемент Arabidopsis , чтобы активировать основные гены ответа на гипоксию в Arabidopsis (33). Наши результаты EMSA расширяют существующие знания, демонстрируя, что фланкирующая последовательность GCC-боксов в промоторах ERF66 и ERF67 также важна и может играть роль в селективности SUB1A-1 для активации транскрипции.
Вместе наши эксперименты (рис. 1 и 2) показывают, что SUB1A-1 непосредственно активирует ERF66 и ERF67 в ответ на погружение посредством взаимодействия с GCC-боксами в их соответствующих промоторных областях.
Сверхэкспрессия ERF66, ERF67 или SUB1A-1 повышает устойчивость трансгенного риса к погружению.
Поскольку ERF66 и ERF67 являются нижележащими целями SUB1A-1, мы затем проверили, участвуют ли ERF66 и ERF67 в устойчивости к погружению.Мы индивидуально сверхэкспрессировали каждый ген ( SI Приложение , рис. S5) в чувствительном к погружению сорте TNG67 (который не содержит SUB1A-1 ) и оценили жизнеспособность полученных трансгенных линий через 7 дней погружения. Используя две независимые линии для каждого трансгена, мы обнаружили, что все три фактора транскрипции, включая SUB1A-1, индивидуально приводили к повышенной устойчивости к погружению по сравнению с WT (рис. 3). В отличие от Xu et al. (5), мы не наблюдали фенотип полукарликов в линиях со сверхэкспрессией TNG67 SUB1A-1 (SUB1A-1 OE), что может быть связано с использованием разных генетических фонов в двух исследованиях.
Рис. 3.
Фенотипы линий сверхэкспрессии SUB1A-1, ERF67 и ERF66 после погружения.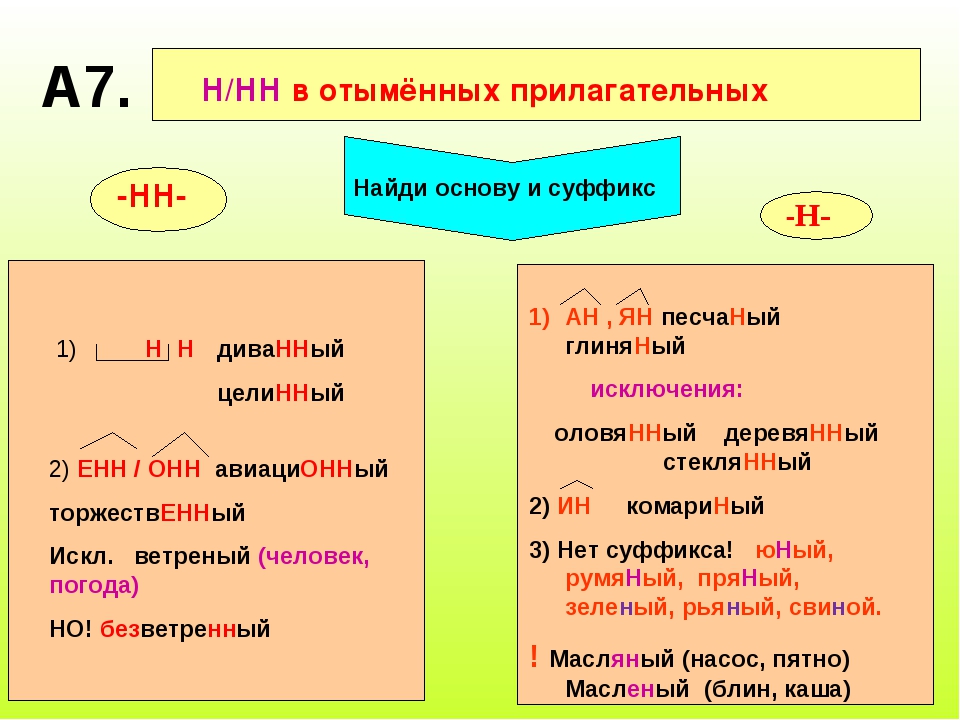 ( A ) Растения риса после 14 дней восстановления после погружения в воду. Четырнадцатидневные растения риса помещали на 7 дней в темноту. После погружения растения возвращали в нормальные условия роста на 14 дней восстановления и фотографировали. ( B ) Жизнеспособность целых растений после удаления всходов. Жизнеспособность всего растения каждого генотипа оценивали в образцах, представленных в A .Растения оценивали как жизнеспособные, если в период восстановления появлялся новый лист. Данные представляют собой средние значения ± стандартное отклонение для двух независимых повторов (* P <0,05 и ** P <0,01 указывают на значимые различия по критерию Стьюдента t ). Значение P в строке 1 ERF67 OE составляет 0,06.
( A ) Растения риса после 14 дней восстановления после погружения в воду. Четырнадцатидневные растения риса помещали на 7 дней в темноту. После погружения растения возвращали в нормальные условия роста на 14 дней восстановления и фотографировали. ( B ) Жизнеспособность целых растений после удаления всходов. Жизнеспособность всего растения каждого генотипа оценивали в образцах, представленных в A .Растения оценивали как жизнеспособные, если в период восстановления появлялся новый лист. Данные представляют собой средние значения ± стандартное отклонение для двух независимых повторов (* P <0,05 и ** P <0,01 указывают на значимые различия по критерию Стьюдента t ). Значение P в строке 1 ERF67 OE составляет 0,06.
ERF66 и ERF67 подпадают под действие правила N-End.
SUB1A-1, ERF66 и ERF67 все имеют консервативную последовательность N-degron MCGG (Рис.1 B и SI Приложение , Рис.S6), но SUB1A-1 не подвергается регуляции правила N-конца in vitro (19). Затем мы исследовали, являются ли ERF66 и ERF67 мишенями пути правила N-конца. Используя ранее установленный анализ in vitro (19), согласно которому белки экспрессируются в системе ретикулоцитов кролика, содержащей консервативные компоненты правила N-конца, мы показали, что мутация цистеина в аланин в положении остатка 2 (C2A) в ERF66-MYC и ERF67-MYC привели к повышению стабильности белка в присутствии ингибитора синтеза белка циклогексимида (CHX) по сравнению с WT, тогда как варианты SUB1A-1-HA WT и C2A не показали разницы в стабильности белка (рис.4 А ). Поскольку N-концевой цистеин имеет решающее значение для обновления ERFVII Arabidopsis (19, 20), эти данные in vitro предполагают, что ERF66 и ERF67, в отличие от SUB1A-1, являются субстратами пути правила N-конца.
Затем мы исследовали, являются ли ERF66 и ERF67 мишенями пути правила N-конца. Используя ранее установленный анализ in vitro (19), согласно которому белки экспрессируются в системе ретикулоцитов кролика, содержащей консервативные компоненты правила N-конца, мы показали, что мутация цистеина в аланин в положении остатка 2 (C2A) в ERF66-MYC и ERF67-MYC привели к повышению стабильности белка в присутствии ингибитора синтеза белка циклогексимида (CHX) по сравнению с WT, тогда как варианты SUB1A-1-HA WT и C2A не показали разницы в стабильности белка (рис.4 А ). Поскольку N-концевой цистеин имеет решающее значение для обновления ERFVII Arabidopsis (19, 20), эти данные in vitro предполагают, что ERF66 и ERF67, в отличие от SUB1A-1, являются субстратами пути правила N-конца.
Рис. 4.
ERF66 и ERF67 являются субстратами пути правила N-конца. ( A ) Анализ in vitro стабильности белка HA- или MYC-меченых вариантов WT и C2A SUB1A-1, SUB1A-2, ERF66 и ERF67 после обработки CHX.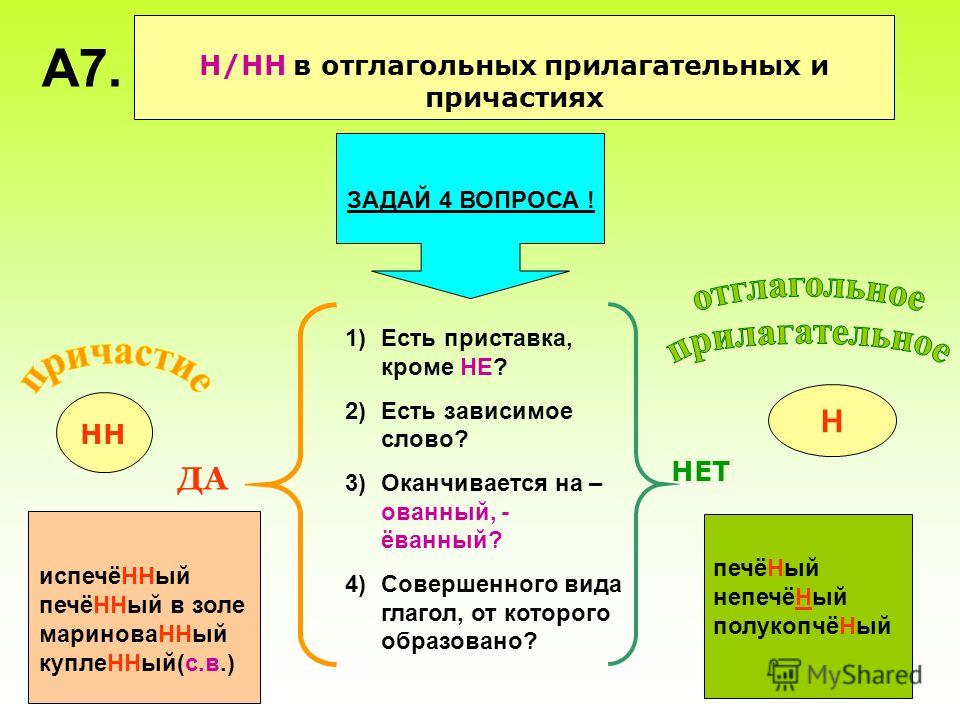 ( B ) Стабильность ERF66 и ERF67, экспрессируемых в протопластах риса, повышается за счет мутации C2A или обработки MG132.Конструкции UbiP :: ERFVII-Luc котрансфицировали в протопласты риса TNG67 с плазмидой UbiP :: GUS , которую использовали в качестве стабильного контроля. Трансфицированные протопласты инкубировали в растворе W5 в течение 3 или 4 часов, а затем собирали для вестерн-блот-анализа ( SI Приложение , фиг. S7 A ), и относительные уровни Luc и GUS были нормализованы по тубулину, соответственно. NT, нетрансфицированный. Молекулярные массы слитых белков ERF66 / 67-Luc составляют ~ 87/85 кДа соответственно.( C и D ) CHX преследует ERF66 и ERF67 с / без MG132 в протопластах риса. Показаны уровни белка ERF66 / ERF67 через 4 часа после трансформации без MG132 (дорожка 1) и с MG132 (дорожка 2). Эксперименты с CHX-преследованием были инициированы после 4-часовой обработки MG132 для обеспечения высоких уровней белка в начале погони путем замены буфера только на CHX (100 мкМ) или CHX и MG132 (20 мкМ).
( B ) Стабильность ERF66 и ERF67, экспрессируемых в протопластах риса, повышается за счет мутации C2A или обработки MG132.Конструкции UbiP :: ERFVII-Luc котрансфицировали в протопласты риса TNG67 с плазмидой UbiP :: GUS , которую использовали в качестве стабильного контроля. Трансфицированные протопласты инкубировали в растворе W5 в течение 3 или 4 часов, а затем собирали для вестерн-блот-анализа ( SI Приложение , фиг. S7 A ), и относительные уровни Luc и GUS были нормализованы по тубулину, соответственно. NT, нетрансфицированный. Молекулярные массы слитых белков ERF66 / 67-Luc составляют ~ 87/85 кДа соответственно.( C и D ) CHX преследует ERF66 и ERF67 с / без MG132 в протопластах риса. Показаны уровни белка ERF66 / ERF67 через 4 часа после трансформации без MG132 (дорожка 1) и с MG132 (дорожка 2). Эксперименты с CHX-преследованием были инициированы после 4-часовой обработки MG132 для обеспечения высоких уровней белка в начале погони путем замены буфера только на CHX (100 мкМ) или CHX и MG132 (20 мкМ). Относительные уровни Luc были нормализованы по тубулину. Данные представляют собой средние значения ± стандартное отклонение от трех независимых повторов (* P <0.05 указывает на значительную разницу по тесту Стьюдента t ).
Относительные уровни Luc были нормализованы по тубулину. Данные представляют собой средние значения ± стандартное отклонение от трех независимых повторов (* P <0.05 указывает на значительную разницу по тесту Стьюдента t ).
Затем мы исследовали регуляцию стабильности ERF66, ERF67 и SUB1A-1 in vivo путем временной экспрессии мутантных вариантов WT и C2A с С-концом, меченных Luc, управляемых промотором Ubi в клетках протопластов риса TNG67 (рис. B и SI Приложение , Рис. S7 A и B ). Здесь мы смогли обнаружить только низкие уровни WT ERF66 / 67 с помощью вестерн-блоттинга, но мутация C2A или обработка MG132 приводили к усиленному накоплению (рис.4 B — D ). Напротив, варианты SUB1A-1 WT и C2A показали аналогичные уровни накопления ( SI, приложение , рис. S7, C ). После обработки CHX накопленные белки WT ERF66 / 67-Luc со временем разлагались, но демонстрировали повышенную стабильность в присутствии MG132, подтверждая, что наблюдаемые различия в уровнях ERF66 / 67 связаны с их регуляцией через путь деградации протеасом (рис.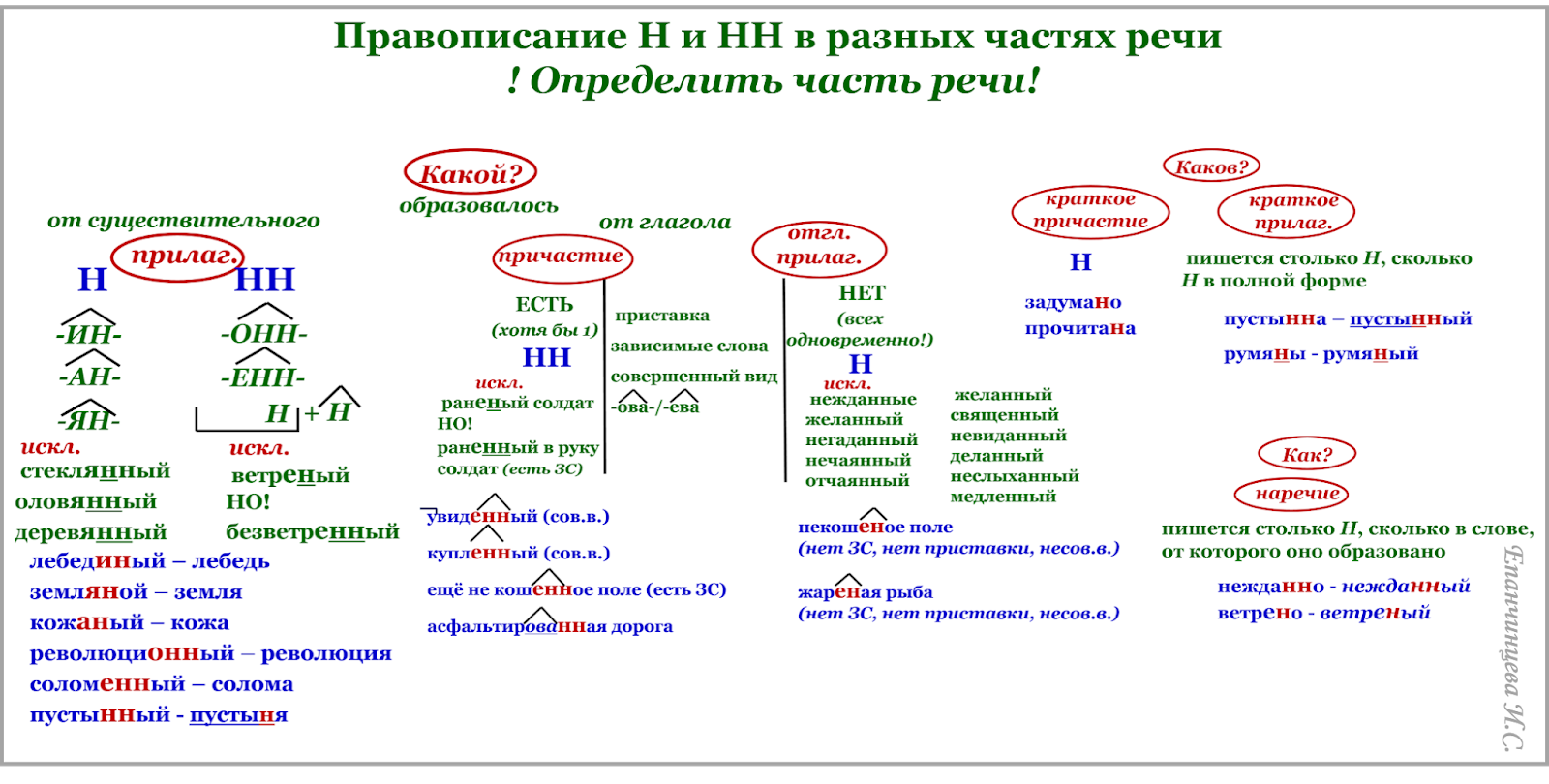 4). C и D ). Мы также исследовали влияние гипоксии, вызванной погружением, на стабильность ERF66 и ERF67 с использованием трансгенного подхода Arabidopsis .Здесь мы наблюдали повышенные уровни белка GFP-меченных ERF66 и ERF67 во время периода погружения ( SI Приложение , рис. S8 A ), тогда как изменение транскрипта оставалось относительно постоянным по сравнению с индуцируемым гипоксией контрольным геном ALCOHOL DEHYDROGENASE1 ( ADh2 ; SI Приложение , рис. S8 B ). Это указывает на то, что гипоксия, вызванная погружением, приводит к стабилизации ERF66 и ERF67, подобно ERFVII Arabidopsis .В совокупности наши анализы стабильности белка показывают, что ERF66 и ERF67 являются субстратами пути правила N-конца, тогда как SUB1A-1 — нет.
4). C и D ). Мы также исследовали влияние гипоксии, вызванной погружением, на стабильность ERF66 и ERF67 с использованием трансгенного подхода Arabidopsis .Здесь мы наблюдали повышенные уровни белка GFP-меченных ERF66 и ERF67 во время периода погружения ( SI Приложение , рис. S8 A ), тогда как изменение транскрипта оставалось относительно постоянным по сравнению с индуцируемым гипоксией контрольным геном ALCOHOL DEHYDROGENASE1 ( ADh2 ; SI Приложение , рис. S8 B ). Это указывает на то, что гипоксия, вызванная погружением, приводит к стабилизации ERF66 и ERF67, подобно ERFVII Arabidopsis .В совокупности наши анализы стабильности белка показывают, что ERF66 и ERF67 являются субстратами пути правила N-конца, тогда как SUB1A-1 — нет.
ERF66 / ERF67 и SUB1A-1 образуют сигнальный каскад для регулирования откликов при погружении в нисходящий поток.
Сообщается, что SUB1A-1 может быть активирован различными абиотическими стрессами. Мы показали, что ERF66 и ERF67 действуют генетически ниже SUB1A-1 и подвергаются регуляции правила N-конца во время гипоксии.Следовательно, мы предположили, что существует два набора генов, один регулируется каскадом SUB1A-1 и ERF66 / ERF67 , а другой регулируется исключительно SUB1A-1. Мы провели анализ секвенирования РНК (RNA-seq) трансгенных линий SUB1A-1 (линия 1), ERF66 (линия 2) и ERF67 (линия 2) (как показано на фиг.3), чтобы изучить эффект сверхэкспрессии этих трансгенных линий. факторы транскрипции на глобальную экспрессию генов (набор данных S1 показывает полные результаты). В этих анализах мы сначала нормализовали уровни экспрессии после 24-часовой обработки погружением с соответствующими уровнями экспрессии при 0-часовой обработке, а затем устранили любые изменения, наблюдаемые в чувствительной к погружению фоновой линии TNG67.При этом мы идентифицировали 271, 285 и 381 гены, которые более чем вдвое активировались в линиях индивидуальной сверхэкспрессии SUB1A-1, ERF67 и ERF66, соответственно, но не в TNG67.
Мы показали, что ERF66 и ERF67 действуют генетически ниже SUB1A-1 и подвергаются регуляции правила N-конца во время гипоксии.Следовательно, мы предположили, что существует два набора генов, один регулируется каскадом SUB1A-1 и ERF66 / ERF67 , а другой регулируется исключительно SUB1A-1. Мы провели анализ секвенирования РНК (RNA-seq) трансгенных линий SUB1A-1 (линия 1), ERF66 (линия 2) и ERF67 (линия 2) (как показано на фиг.3), чтобы изучить эффект сверхэкспрессии этих трансгенных линий. факторы транскрипции на глобальную экспрессию генов (набор данных S1 показывает полные результаты). В этих анализах мы сначала нормализовали уровни экспрессии после 24-часовой обработки погружением с соответствующими уровнями экспрессии при 0-часовой обработке, а затем устранили любые изменения, наблюдаемые в чувствительной к погружению фоновой линии TNG67.При этом мы идентифицировали 271, 285 и 381 гены, которые более чем вдвое активировались в линиях индивидуальной сверхэкспрессии SUB1A-1, ERF67 и ERF66, соответственно, но не в TNG67. Интересно, что ERF66 и ERF67 были приблизительно в 3- и 30-кратно активированы в линии сверхэкспрессии SUB1A-1, соответственно. Кроме того, гены 422, 487 и 511 подавлялись более чем в два раза в индивидуальных линиях сверхэкспрессии SUB1A-1, ERF67 и ERF66 соответственно (рис.5 А ). Анализ списков генов выявил две отдельные группы дифференциально экспрессируемых генов, одна из которых зависит от SUB1A-1 с помощью ERF66 и / или ERF67, а другая регулируется исключительно SUB1A-1 (рис. 5 B и C ). ). Для первой группы анализ диаграммы Венна показывает, что в общей сложности 151 ген был активирован в SUB1A-1 и ERF67 (24 гена), SUB1A-1 и ERF66 (60 генов) и во всех трех отдельных линиях сверхэкспрессии (67 генов), Это указывает на то, что ERF66 и ERF67 могут иметь разные нисходящие цели (рис.5 В ). Между тем, мы также обнаружили в общей сложности 217 генов, которые подавлялись в SUB1A-1 и ERF67 (54 гена), SUB1A-1 и ERF66 (56 генов), а также во всех трех отдельных линиях сверхэкспрессии (107 генов; рис.
Интересно, что ERF66 и ERF67 были приблизительно в 3- и 30-кратно активированы в линии сверхэкспрессии SUB1A-1, соответственно. Кроме того, гены 422, 487 и 511 подавлялись более чем в два раза в индивидуальных линиях сверхэкспрессии SUB1A-1, ERF67 и ERF66 соответственно (рис.5 А ). Анализ списков генов выявил две отдельные группы дифференциально экспрессируемых генов, одна из которых зависит от SUB1A-1 с помощью ERF66 и / или ERF67, а другая регулируется исключительно SUB1A-1 (рис. 5 B и C ). ). Для первой группы анализ диаграммы Венна показывает, что в общей сложности 151 ген был активирован в SUB1A-1 и ERF67 (24 гена), SUB1A-1 и ERF66 (60 генов) и во всех трех отдельных линиях сверхэкспрессии (67 генов), Это указывает на то, что ERF66 и ERF67 могут иметь разные нисходящие цели (рис.5 В ). Между тем, мы также обнаружили в общей сложности 217 генов, которые подавлялись в SUB1A-1 и ERF67 (54 гена), SUB1A-1 и ERF66 (56 генов), а также во всех трех отдельных линиях сверхэкспрессии (107 генов; рис. 5 C). ).
5 C). ).
Рис. 5.
Транскриптомный анализ линий сверхэкспрессии SUB1A-1, ERF66 и ERF67 при погружении. ( A ) Гистограмма, показывающая количество генов с повышенной и отрицательной регуляцией (более чем в два раза; P <0,05) в линиях сверхэкспрессии SUB1A-1 / ERF67 / ERF66, которые не регулируются дифференцированно в TNG67.( B ) Перекрытие между генами, значительно активируемое сверхэкспрессией SUB1A-1, ERF66 и ERF66 и распределением функциональных категорий активированных генов. ( C ) Перекрытие между генами, значительно подавляемыми SUB1A-1, ERF66 и ERF67, и распределение функциональных категорий генов с подавленной экспрессией. Гистограммы в C и D указывают значения P расширенных функциональных категорий. ( D ) Репрезентативные гены активируются в линиях сверхэкспрессии SUB1A-1 и ERF66 / 67.Первые семь генов являются ортологами основных генов гипоксии, которые активируются в линиях сверхэкспрессии SUB1A-1 и ERF66 / 67.
Анализ онтологии генов показал, что гены с повышенной регуляцией участвуют в различных процессах, включая ответ на стресс, защитный ответ, фосфорилирование и активность протеинкиназы (рис. 5 B ), тогда как гены с пониженной регуляцией включали гены, связанные с углеводный обмен и клеточный липидный обмен (рис. 5 C ).Активно регулируемые гены во всех трех трансгенных линиях включали ортологи основных генов, связанных с гипоксией, включая несимбиотический гемоглобин 1/2 (LOC_Os03g13140 / LOC_Os03g12510), алкогольдегидрогеназу 1/2 (LOC_Os11g10480 / LOC_Os11g10510). LOC_Os08g03420), Dreg-2-подобный протеин / галогенкислоту, дегалогеназоподобный гидролазный домен, содержащий белок 3 (LOC_Os02g07730), и фосфофруктокиназу 5 (LOC_Os05g44922; Фиг.5 D и Dataset S1). Это обеспечивает транскрипционное объяснение повышенной устойчивости этих линий сверхэкспрессии к погружению (рис.3), и дополнительно подтверждает участие всех трех ERFVII в координации откликов при погружении.
N-конец SUB1A-1 имеет случайную структуру катушки.
Ключевые ферменты в пути правила N-конца, включая MetAP, ATE и PRT6, высоко консервативны у эукариот (22, 23). Активный сайт связывания функционального гомолога PRT6 человека, UBR1, представляет собой неглубокий и в основном гидрофобный карман, в который могут поместиться активированные N-дегроны (34, 35). Чтобы понять, как SUB1A-1 может уклоняться от регуляции правила N-конца, несмотря на наличие N-концевого мотива, аналогичного At ERFVII и Os ERF66 / 67, мы использовали КД и ЯМР-спектроскопию для исследования рекомбинантных SUB1A-1 и SUB1A- 1 N-конец (SUB1A-1N), который состоит из первых 115 а.о. SUB1A-1 ( SI Приложение , рис.S9) с дополнительным N-концевым серином (остаток сайта расщепления протеазой TEV) и C-концевой His-меткой для продукции и очистки белка. Исследование вторичной структуры с помощью CD показало, что полноразмерный SUB1A-1 в основном неструктурирован, а SUB1A-1N также напоминает случайную катушку (рис. 6 A и B ). Далее мы проанализировали структурные свойства SUB1A-1N с помощью ЯМР-спектроскопии. Как показано на рис. 6 C , 2D 1 H, 15 N-полосно-селективный коротко-переходный (BEST) -гетероядерный спектр одноквантовой когерентности (HSQC) показывает, что кросс-пики основной цепи NH группы SUB1A-1N встречаются в очень узком диапазоне химического сдвига, что указывает на случайную структуру катушки при объединении этого результата с его кривой CD случайной катушки.Кроме того, экспонированный растворителем спектр амидного протона 2D 1 H, 15 N-HSQC SUB1A-1N показывает, что большинство амидных протонов обменялись кросс-пиками с водой (со скоростью обмена более 3 Гц), что указывает на что амиды основной цепи подвергаются воздействию растворителя и не защищены структурными или водородными связями (рис. 6 D ). Таким образом, комбинированный анализ CD и ЯМР показывает, что SUB1A-1N неструктурирован, что позволяет предположить, что N-конец SUB1A-1 очень гибкий и должен распознаваться компонентами правила N-конца.
6 A и B ). Далее мы проанализировали структурные свойства SUB1A-1N с помощью ЯМР-спектроскопии. Как показано на рис. 6 C , 2D 1 H, 15 N-полосно-селективный коротко-переходный (BEST) -гетероядерный спектр одноквантовой когерентности (HSQC) показывает, что кросс-пики основной цепи NH группы SUB1A-1N встречаются в очень узком диапазоне химического сдвига, что указывает на случайную структуру катушки при объединении этого результата с его кривой CD случайной катушки.Кроме того, экспонированный растворителем спектр амидного протона 2D 1 H, 15 N-HSQC SUB1A-1N показывает, что большинство амидных протонов обменялись кросс-пиками с водой (со скоростью обмена более 3 Гц), что указывает на что амиды основной цепи подвергаются воздействию растворителя и не защищены структурными или водородными связями (рис. 6 D ). Таким образом, комбинированный анализ CD и ЯМР показывает, что SUB1A-1N неструктурирован, что позволяет предположить, что N-конец SUB1A-1 очень гибкий и должен распознаваться компонентами правила N-конца. Это повышает вероятность того, что другие области SUB1A-1 могут участвовать в предотвращении деградации с помощью пути правила N-конца или что другие белки связываются с SUB1A-1, чтобы защитить N-degron.
Это повышает вероятность того, что другие области SUB1A-1 могут участвовать в предотвращении деградации с помощью пути правила N-конца или что другие белки связываются с SUB1A-1, чтобы защитить N-degron.
Рис. 6.
Спектры КД и ЯМР рекомбинантных конструкций SUB1A-1. Спектры КД полноразмерного SUB1A-1 ( A ) и только N-конца SUB1A-1 ( B ) показывают, что SUB1A-1 и его N-конец в основном неструктурированы. Перекрестные пики 2D 1 H, 15 N-BEST-HSQC-спектр конца SUB1A-1 N встречаются только в очень узком диапазоне химического сдвига ( C ), что указывает на случайную структуру катушки, когда он комбинируется с результат CD.Обнаруженный растворителем амидный протон 2D 1 H, 15 N-HSQC спектр только N-конца SUB1A-1 ( D ) показывает, что большая часть амидных протонов имеет кросс-пики обмена с водой, что указывает на то, что амиды являются подвержены воздействию растворителей.
Конечный конец C SUB1A-1 предотвращает его деградацию из-за пути правила N-конца.

Чтобы проверить, могут ли другие области SUB1A-1 влиять на его деградацию по правилу N-конца, мы проанализировали стабильность белка двух усеченных на С-конце вариантов SUB1A-1 в протопласте риса TNG67, включая ( i ) СУБ1А-1Н (т.е., только N-конец SUB1A-1) и ( ii ) SUB1A-1ΔC (SUB1A-1 без С-конца; SI Приложение , Рис. S7 B ). Уровни белка SUB1A-1N и SUB1A-1ΔC были аналогичны WT ERF67, но значительно ниже, чем ERF67 (C2A), что указывает на то, что усеченный SUB1A-1 нестабилен после удаления С-конца (фиг.7 A и B ) . Чтобы подтвердить, что эта нестабильность вызвана деградацией через путь правила N-конца, мы временно экспрессировали варианты C2A ( SI Приложение , рис.S7 B ) обеих укороченных конструкций в клетках протопластов риса TNG67. Количество белка SUB1A-1 (C2A) N и SUB1A-1 (C2A) ΔC было намного выше, чем SUB1A-1N и SUB1A-1ΔC, и аналогично ERF67 (C2A) (фиг.7 A и B ). Это предполагает, что в отличие от полноразмерного SUB1A-1 ( SI Приложение , рис. S7, C ), усеченные на C-конце варианты SUB1A-1 деградируют посредством пути правила N-конца. Чтобы понять, как С-конец SUB1A-1 вмешивается в деградацию SUB1A-1, мы исследовали способность этих двух областей SUB1A-1 взаимодействовать друг с другом.Двухгибридный анализ дрожжей выявил взаимодействие между N-концом SUB1A-1 и С-концом (фиг. 7 C ). Это было специфично для С-конца SUB1A-1, поскольку С-концы ERF66 и ERF67 не взаимодействовали с N-концом SUB1A-1. Это взаимодействие было также подтверждено экспериментами изотермической калориметрии титрования (iTC) с использованием рекомбинантного N-конца и С-конца SUB1A-1 (фиг. 7 D ). Таким образом, мы предполагаем, что C-концевая область SUB1A физически взаимодействует с N-концом SUB1A-1, и что это экранирует N-degron, предотвращая обмен белка.
S7, C ), усеченные на C-конце варианты SUB1A-1 деградируют посредством пути правила N-конца. Чтобы понять, как С-конец SUB1A-1 вмешивается в деградацию SUB1A-1, мы исследовали способность этих двух областей SUB1A-1 взаимодействовать друг с другом.Двухгибридный анализ дрожжей выявил взаимодействие между N-концом SUB1A-1 и С-концом (фиг. 7 C ). Это было специфично для С-конца SUB1A-1, поскольку С-концы ERF66 и ERF67 не взаимодействовали с N-концом SUB1A-1. Это взаимодействие было также подтверждено экспериментами изотермической калориметрии титрования (iTC) с использованием рекомбинантного N-конца и С-конца SUB1A-1 (фиг. 7 D ). Таким образом, мы предполагаем, что C-концевая область SUB1A физически взаимодействует с N-концом SUB1A-1, и что это экранирует N-degron, предотвращая обмен белка.
Рис. 7.
Усеченный на C-конце SUB1A-1 может быть ухудшен с помощью пути правила N-конца. Вестерн-блоттинг-анализ стабильности белка усеченного SUB1A-1 в протопластах риса TNG67. ( A и B ) Анализы стабильности белков SUB1A-1N и SUB1A-1ΔC соответственно. Конструкции UbiP :: SUB1A-1N / SUB1A-1ΔC / ERF67-Luc котрансфицировали в протопласты риса TNG67 с плазмидой UbiP :: GUS , которую использовали в качестве стабильного контроля. Трансфицированные протопласты инкубировали в растворе W5 в течение 3 или 4 часов, а затем собирали для дальнейшего вестерн-блоттинга, и относительные уровни Luc и GUS нормализовали по тубулину, соответственно.NT, нетрансфицированный. Данные представляют собой средние значения ± стандартное отклонение для трех независимых повторов (* P <0,05 указывает на значительную разницу по критерию Стьюдента t ). Молекулярная масса слитых белков SUB1A-1N / SUB1A-1ΔC-Luc составляет ∼74 / 81 кДа соответственно. Молекулярная масса слитого белка ERF67-Luc составляет ~ 85 кДа. ( C ) Дрожжевой двугибридный анализ тестирует взаимодействия между N-концом SUB1A-1 и C-концом SUB1A-1, ERF66 или ERF67.
( A и B ) Анализы стабильности белков SUB1A-1N и SUB1A-1ΔC соответственно. Конструкции UbiP :: SUB1A-1N / SUB1A-1ΔC / ERF67-Luc котрансфицировали в протопласты риса TNG67 с плазмидой UbiP :: GUS , которую использовали в качестве стабильного контроля. Трансфицированные протопласты инкубировали в растворе W5 в течение 3 или 4 часов, а затем собирали для дальнейшего вестерн-блоттинга, и относительные уровни Luc и GUS нормализовали по тубулину, соответственно.NT, нетрансфицированный. Данные представляют собой средние значения ± стандартное отклонение для трех независимых повторов (* P <0,05 указывает на значительную разницу по критерию Стьюдента t ). Молекулярная масса слитых белков SUB1A-1N / SUB1A-1ΔC-Luc составляет ∼74 / 81 кДа соответственно. Молекулярная масса слитого белка ERF67-Luc составляет ~ 85 кДа. ( C ) Дрожжевой двугибридный анализ тестирует взаимодействия между N-концом SUB1A-1 и C-концом SUB1A-1, ERF66 или ERF67. AD, домен активации GAL4; BD, ДНК-связывающий домен GAL4.Положительные взаимодействия представлены ростом на среде с тройным отсевом (-WLH), которая проверяет экспрессию репортерного гена HIS3 . Рост дрожжей на среде с двойным выпадением (-WL) включен в качестве контроля котрансформации. ( D ) iTC эксперимент по связыванию между SUB1A-1 N- и C-концами. Верхняя кривая показывает скорректированные тепловые импульсы, полученные в результате титрования концевой заделки SUB1A-1 C, а нижняя диаграмма показывает интегрированный тепловой импульс вместе с подгонкой.
AD, домен активации GAL4; BD, ДНК-связывающий домен GAL4.Положительные взаимодействия представлены ростом на среде с тройным отсевом (-WLH), которая проверяет экспрессию репортерного гена HIS3 . Рост дрожжей на среде с двойным выпадением (-WL) включен в качестве контроля котрансформации. ( D ) iTC эксперимент по связыванию между SUB1A-1 N- и C-концами. Верхняя кривая показывает скорректированные тепловые импульсы, полученные в результате титрования концевой заделки SUB1A-1 C, а нижняя диаграмма показывает интегрированный тепловой импульс вместе с подгонкой.
Обсуждение
Факторы транскрипции ERFVII участвуют в чувствительных к гипоксии и регулирующих ответах на наводнение и / или гипоксический стресс.Например, все пять ERFVII в Arabidopsis функционируют как важные регуляторы устойчивости к затоплению и / или гипоксии, а ERFVII у ячменя, Rumex и Rorippa регулируют реакцию на заболачивание (12, 15, 17, 29, 36). № – 40).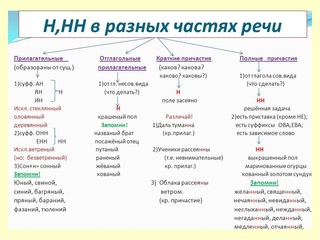 Кроме того, ERFVII Arabidopsis также были связаны с другими абиотическими и биотическими ответами (12, 41–43). У риса ERFVII SUB1A-1 является главным регулятором реакции покоя погружение-выживание, а также других абиотических стрессов (27, 28).Однако, в отличие от всех других исследованных ERFVII, было показано, что SUB1A-1 сопротивляется пути правила N-конца, указывая тем самым, что он не принимает непосредственного участия в восприятии гипоксии. Кроме того, SUB1A-1 не придает устойчивость к затоплению у арабидопсиса , что предполагает некоторую степень различия между рисом и механизмом покоя арабидопсиса в ответ на стресс погружения. Здесь мы предлагаем регуляторный каскад в SUB1A-1 -зависимой реакции погружения, который включает два других риса ERFVII , ERF66 и ERF67 .
Кроме того, ERFVII Arabidopsis также были связаны с другими абиотическими и биотическими ответами (12, 41–43). У риса ERFVII SUB1A-1 является главным регулятором реакции покоя погружение-выживание, а также других абиотических стрессов (27, 28).Однако, в отличие от всех других исследованных ERFVII, было показано, что SUB1A-1 сопротивляется пути правила N-конца, указывая тем самым, что он не принимает непосредственного участия в восприятии гипоксии. Кроме того, SUB1A-1 не придает устойчивость к затоплению у арабидопсиса , что предполагает некоторую степень различия между рисом и механизмом покоя арабидопсиса в ответ на стресс погружения. Здесь мы предлагаем регуляторный каскад в SUB1A-1 -зависимой реакции погружения, который включает два других риса ERFVII , ERF66 и ERF67 .
Мы проанализировали кинетику транскрипции 16 OsERFVII s у двух сортов indica (устойчивый к погружению FR13A и чувствительный к погружению IR29) и обнаружили, что многие из них транскрипционно активируются в ответ на обработку погружением ( SI Приложение , Рис. S1). Сюда входит ERF70, который, как недавно было показано, способствует лучшему восстановлению после стресса при погружении (44). Пять из этих OsERFVII ( ERF59 , ERF60 , ERF61 , ERF66 и ERF67 ) имели более высокие уровни транскриптов в FR13A, чем в IR29, и их паттерны экспрессии были аналогичны таковым у SUB1A. 1 в FR13A и IR29 ( SI Приложение , рис.S1). Путем перекрестного исследования уровней транскриптов этих пяти ERFVII в других устойчивых к погружению сортах IR64 ( Sub1 ) и Swarna ( Sub1 ) и других чувствительных сортах, IR64 и Swarna, мы обнаружили, что только ERF66 и ERF67 показали повышенное количество транскриптов в толерантных сортах, чем в чувствительных сортах (фиг. 1), указывая на то, что они являются нижележащими мишенями для SUB1A-1 во время погружения.
S1). Сюда входит ERF70, который, как недавно было показано, способствует лучшему восстановлению после стресса при погружении (44). Пять из этих OsERFVII ( ERF59 , ERF60 , ERF61 , ERF66 и ERF67 ) имели более высокие уровни транскриптов в FR13A, чем в IR29, и их паттерны экспрессии были аналогичны таковым у SUB1A. 1 в FR13A и IR29 ( SI Приложение , рис.S1). Путем перекрестного исследования уровней транскриптов этих пяти ERFVII в других устойчивых к погружению сортах IR64 ( Sub1 ) и Swarna ( Sub1 ) и других чувствительных сортах, IR64 и Swarna, мы обнаружили, что только ERF66 и ERF67 показали повышенное количество транскриптов в толерантных сортах, чем в чувствительных сортах (фиг. 1), указывая на то, что они являются нижележащими мишенями для SUB1A-1 во время погружения.
Используя анализы активации транс , мы обнаружили, что SUB1A-1 может транскрипционно активировать ERF66 и ERF67 (рис. 2 A и B ). Мы также подтвердили, что SUB1A-1 может взаимодействовать с промотором ERF66 и ERF67 с помощью ChIP-qPCR (фиг. 2 C ). Идентифицировано несколько боксов GCC с разными фланкирующими последовательностями в промоторных областях ERF66 и ERF67 ( SI Приложение , рис. S4 B ), и наши исследования EMSA показали, что рекомбинантный SUB1A-1 избирательно связывается с несколькими ( но не все) идентифицированных боксов GCC (рис.2 D и SI Приложение , Рис. S4 C ). В совокупности наши данные предполагают, что SUB1A-1 напрямую активирует ERF66 и ERF67 посредством взаимодействия с GCC-боксами в их промоторах, и ERF66 и ERF67 , следовательно, являются нижестоящими мишенями для SUB1A-1. Более того, сверхэкспрессия ERF66 или ERF67 в чувствительном к погружению сорте TNG67 привела к повышенной устойчивости к погружению (рис. 3).
2 A и B ). Мы также подтвердили, что SUB1A-1 может взаимодействовать с промотором ERF66 и ERF67 с помощью ChIP-qPCR (фиг. 2 C ). Идентифицировано несколько боксов GCC с разными фланкирующими последовательностями в промоторных областях ERF66 и ERF67 ( SI Приложение , рис. S4 B ), и наши исследования EMSA показали, что рекомбинантный SUB1A-1 избирательно связывается с несколькими ( но не все) идентифицированных боксов GCC (рис.2 D и SI Приложение , Рис. S4 C ). В совокупности наши данные предполагают, что SUB1A-1 напрямую активирует ERF66 и ERF67 посредством взаимодействия с GCC-боксами в их промоторах, и ERF66 и ERF67 , следовательно, являются нижестоящими мишенями для SUB1A-1. Более того, сверхэкспрессия ERF66 или ERF67 в чувствительном к погружению сорте TNG67 привела к повышенной устойчивости к погружению (рис. 3).
Проведя исследования стабильности белков (рис. 4 и SI Приложение , рис. S7 и S8), мы показали, что ERF66 и ERF67, но не SUB1A-1 (19), являются субстратами для пути правила N-конца, несмотря на то, что все три белка имеют канонические последовательности N-degron на своих N-концах. Анализ ЯМР показал, что N-конец SUB1A-1 представляет собой структуру случайной спирали (фиг. 6), что указывает на то, что N-конец SUB1A-1 очень гибкий и должен легко распознаваться ферментами, связанными с правилом N-конца. Это подняло вопрос о том, как SUB1A-1 может избежать регулирования правила N-конца.
4 и SI Приложение , рис. S7 и S8), мы показали, что ERF66 и ERF67, но не SUB1A-1 (19), являются субстратами для пути правила N-конца, несмотря на то, что все три белка имеют канонические последовательности N-degron на своих N-концах. Анализ ЯМР показал, что N-конец SUB1A-1 представляет собой структуру случайной спирали (фиг. 6), что указывает на то, что N-конец SUB1A-1 очень гибкий и должен легко распознаваться ферментами, связанными с правилом N-конца. Это подняло вопрос о том, как SUB1A-1 может избежать регулирования правила N-конца.
Наши анализы (рис. 7 A и B ) показали, что усеченный на С-конце SUB1A-1 может разрушаться через путь правила N-конца, предполагая, что С-конец SUB1A-1 участвует в ингибировании его деградации. . Эксперименты с двугибридными дрожжами и iTC показали физическое взаимодействие между N- и C-концами SUB1A-1 (фиг. 7 C и D ). Следовательно, вероятно, что С-конец SUB1A-1 помогает маскировать N-концевую область, участвующую в пути правила N-конца. Аналогичный сценарий описан для белка α-синуклеина, при котором дальнодействующие междоменные взаимодействия приводят к стабилизации за счет принятия ансамбля конформаций для маскировки его амилоидогенного домена (45, 46). Взятые вместе, эти результаты предполагают, что особенности на N-конце и C-конце SUB1A-1 способствуют его уходу от деградации правила N-конца, вероятно, за счет доменных взаимодействий, которые предотвращают адекватное воздействие на N-конец или блокируют сайт убиквитинирования. . Однако подробный молекулярный механизм остается неясным и требует дальнейшего изучения.
Аналогичный сценарий описан для белка α-синуклеина, при котором дальнодействующие междоменные взаимодействия приводят к стабилизации за счет принятия ансамбля конформаций для маскировки его амилоидогенного домена (45, 46). Взятые вместе, эти результаты предполагают, что особенности на N-конце и C-конце SUB1A-1 способствуют его уходу от деградации правила N-конца, вероятно, за счет доменных взаимодействий, которые предотвращают адекватное воздействие на N-конец или блокируют сайт убиквитинирования. . Однако подробный молекулярный механизм остается неясным и требует дальнейшего изучения.
SUB1A-1 является основным фактором, определяющим устойчивость риса к погружению в воду. Он активируется не только при погружении, но и во время засухи, продолжительного темноты, окислительного стресса и этиленового стресса, и играет ключевую роль в ряде реакций на абиотический стресс в дополнение к погружению (8, 27, 28, 47, 48). Отделение SUB1A-1 от регуляции правила N-конца, возможно, позволило SUB1A-1 принять более широкий спектр функций в качестве главного регулятора транскрипции, функционирующего в качестве концентратора для организации сигнальных сетей в ответ на различные стрессы в нормоксических или гипоксических условиях. .Однако это вызвало вопрос о том, как рис отличает стресс погружения (гипоксию) от других регулируемых напряжений SUB1A-1 , которые возникают, когда кислород легко доступен. В этом исследовании мы идентифицировали ERF66 и ERF67 как прямые нижестоящие мишени для SUB1A-1 и субстратов пути правила N-конца, которые могут иметь решающее значение для координации ответов на гипоксию. Кроме того, анализ РНК-seq показал, что две разные группы генов были индуцированы SUB1A-1; одна группа зависит от ERF66 и ERF67, а другая группа не зависит от ERF66 и ERF67 (рис.5 и набор данных S1). SUB1A-1 представляет собой фактор транскрипции, участвующий в нескольких важных процессах при подводном стрессе (49). Наши данные предполагают, что ERF66 и ERF67 являются нижележащими генами SUB1A-1, и эти два гена участвуют в нескольких важных процессах во время погружения и придают устойчивость рису к погружению, а также SUB1A-1 . Это похоже на ситуацию в Arabidopsis , в котором ERFVII HRE1 и HRE2 находятся ниже RAP2.
.Однако это вызвало вопрос о том, как рис отличает стресс погружения (гипоксию) от других регулируемых напряжений SUB1A-1 , которые возникают, когда кислород легко доступен. В этом исследовании мы идентифицировали ERF66 и ERF67 как прямые нижестоящие мишени для SUB1A-1 и субстратов пути правила N-конца, которые могут иметь решающее значение для координации ответов на гипоксию. Кроме того, анализ РНК-seq показал, что две разные группы генов были индуцированы SUB1A-1; одна группа зависит от ERF66 и ERF67, а другая группа не зависит от ERF66 и ERF67 (рис.5 и набор данных S1). SUB1A-1 представляет собой фактор транскрипции, участвующий в нескольких важных процессах при подводном стрессе (49). Наши данные предполагают, что ERF66 и ERF67 являются нижележащими генами SUB1A-1, и эти два гена участвуют в нескольких важных процессах во время погружения и придают устойчивость рису к погружению, а также SUB1A-1 . Это похоже на ситуацию в Arabidopsis , в котором ERFVII HRE1 и HRE2 находятся ниже RAP2. 2, 2.3 и 2.12 (15). Мы предполагаем, что этот регуляторный каскад SUB1A-1 –to– ERF66 / ERF67 является связующим звеном, которое позволяет рису различать погружение в воду и другие абиотические стрессы (рис. 8). В этой модели SUB1A-1 индуцируется при различных абиотических стрессах, что, в свою очередь, активирует гены ERF66 / ERF67 и набор общих генов ответа на стресс. В условиях нормоксического абиотического стресса ERF66 и ERF67 деградируют посредством пути правила N-конца.Только в условиях низкого содержания кислорода ERF66 и ERF67 могут стабилизироваться, накапливаться, чтобы вызвать гипоксические реакции, и позволить FR13A и устойчивым к затоплению сортам выжить до 2 недель при полном погружении. Конститутивная стабильность SUB1A-1 означает, что, когда уровни кислорода вернутся к норме после десорбции, ERF66 / 67 будет быстро деградировать, чтобы выключить специфический транскрипционный ответ гипоксии, но SUB1A-1 останется стабильным для координации экспрессии других генов.
2, 2.3 и 2.12 (15). Мы предполагаем, что этот регуляторный каскад SUB1A-1 –to– ERF66 / ERF67 является связующим звеном, которое позволяет рису различать погружение в воду и другие абиотические стрессы (рис. 8). В этой модели SUB1A-1 индуцируется при различных абиотических стрессах, что, в свою очередь, активирует гены ERF66 / ERF67 и набор общих генов ответа на стресс. В условиях нормоксического абиотического стресса ERF66 и ERF67 деградируют посредством пути правила N-конца.Только в условиях низкого содержания кислорода ERF66 и ERF67 могут стабилизироваться, накапливаться, чтобы вызвать гипоксические реакции, и позволить FR13A и устойчивым к затоплению сортам выжить до 2 недель при полном погружении. Конститутивная стабильность SUB1A-1 означает, что, когда уровни кислорода вернутся к норме после десорбции, ERF66 / 67 будет быстро деградировать, чтобы выключить специфический транскрипционный ответ гипоксии, но SUB1A-1 останется стабильным для координации экспрессии других генов. которые необходимы для десорбции / засухи / выживания АФК.
которые необходимы для десорбции / засухи / выживания АФК.
Рис. 8. Модель
регуляторного каскада SUB1A-1 , ERF66 и ERF67 , который включает пути транскрипции и правила N-конца в ответ на стресс погружения у устойчивых к погружению сортов риса.
Материалы и методы
Более подробная информация об экспериментальных процедурах представлена в Приложении SI , SI Материалы и методы .
Растительные материалы.
Рис ( O. sativa ) сортов FR13A, IR29, IR64, Swarna и TNG67.Две почти изогенные линии, IR64 ( Sub1 ) и Swarna ( Sub1 ), были предоставлены Национальным центром генетических ресурсов растений Тайваньского института сельскохозяйственных исследований (Тайчжун, Тайвань). Трансгенные линии риса со сверхэкспрессией SUB1A-1 / ERF66 / ERF67 (SUB1A-1 OE / ERF66 OE / ERF67 OE) были получены путем преобразования UbiP :: GST-SUB1A-1 / ERF66 / ERF67 в векторе pCAMBIA1301 в TNG67 рис. Трансгенные линии Arabidopsis со сверхэкспрессией ERF66 и ERF67 были получены путем трансформации 35S :: ERF66 / ERF67 — GFP в векторе pK7FWG2 в Arabidopsis ( A . thaliana ) экотип Колумбия-0. Трансформацию в Agrobacterium tumefaciens и Arabidopsis проводили согласно установленным протоколам (50). Трансформация риса была проведена основной лабораторией трансгенных растений Academia Sinica.
Трансгенные линии Arabidopsis со сверхэкспрессией ERF66 и ERF67 были получены путем трансформации 35S :: ERF66 / ERF67 — GFP в векторе pK7FWG2 в Arabidopsis ( A . thaliana ) экотип Колумбия-0. Трансформацию в Agrobacterium tumefaciens и Arabidopsis проводили согласно установленным протоколам (50). Трансформация риса была проведена основной лабораторией трансгенных растений Academia Sinica.
Условия выращивания и обработка погружением.
Семена риса стерилизовали 1,2% (об. / Об.) Гипохлоритом натрия, содержащим 0,1% (об. / Об.) Твин 20, в течение 30 минут и промывали не менее пяти раз стерильной водой.Стерилизованные семена помещали на влажную фильтровальную бумагу в чашки Петри при 37 ° C в темноте на 4 дня. После инкубации равномерно проросшие семена пересаживали на железную сетку в стакан с четвертью крепости раствором Kimura B, pH 5,6–5,8 (51), и раствор обновляли каждые 2 дня.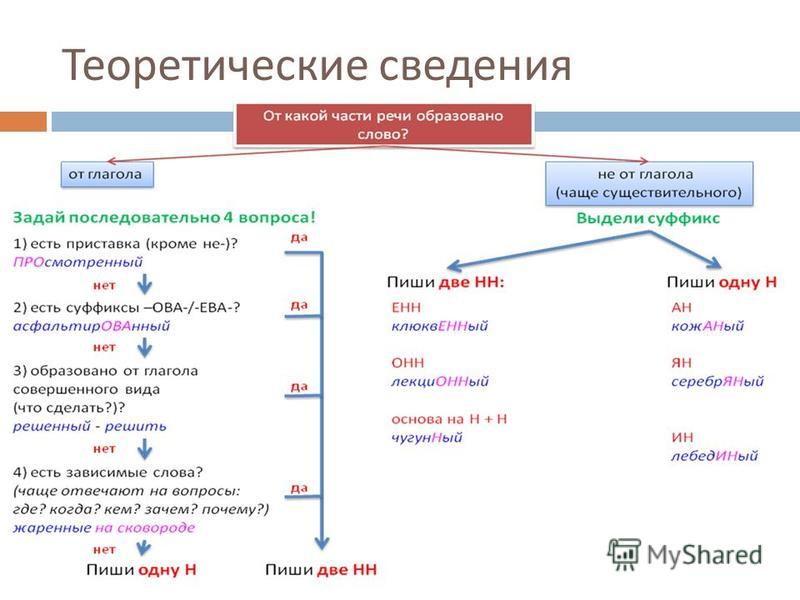 Гидропонно культивируемые проростки выращивали в камере для выращивания при 28 ° C с циклом 16 часов света (120–125 мкмоль μм –2 6s –1 ) / 8 часов темноты до достижения им возраста 14 дней. Для обработки под водой 14-дневных проростков риса химические стаканы с растениями помещали в резервуар для воды (ширина 40 см, длина 40 см, высота 70 см), заполненный водопроводной водой высотой 55 см, на указанное время при температуре 28 ° C в темнота.Для фенотипических анализов данные были собраны для каждого генотипа в двух независимых экспериментах. Четырнадцатидневные проростки подвергали обработке погружением, как описано ранее, в течение 7 дней в темноте. После обработки погружением проростки риса помещали обратно в камеру для выращивания при 28 ° C с циклом 16 часов света (120–125 мкмоль · м −2 s −1 ) / 8 часов темноты для дальнейшего использования. 14 дней восстановления с последующей оценкой жизнеспособности всего растения. Растения оценивали как жизнеспособные, когда в период восстановления появлялся один или несколько новых листьев.
Гидропонно культивируемые проростки выращивали в камере для выращивания при 28 ° C с циклом 16 часов света (120–125 мкмоль μм –2 6s –1 ) / 8 часов темноты до достижения им возраста 14 дней. Для обработки под водой 14-дневных проростков риса химические стаканы с растениями помещали в резервуар для воды (ширина 40 см, длина 40 см, высота 70 см), заполненный водопроводной водой высотой 55 см, на указанное время при температуре 28 ° C в темнота.Для фенотипических анализов данные были собраны для каждого генотипа в двух независимых экспериментах. Четырнадцатидневные проростки подвергали обработке погружением, как описано ранее, в течение 7 дней в темноте. После обработки погружением проростки риса помещали обратно в камеру для выращивания при 28 ° C с циклом 16 часов света (120–125 мкмоль · м −2 s −1 ) / 8 часов темноты для дальнейшего использования. 14 дней восстановления с последующей оценкой жизнеспособности всего растения. Растения оценивали как жизнеспособные, когда в период восстановления появлялся один или несколько новых листьев.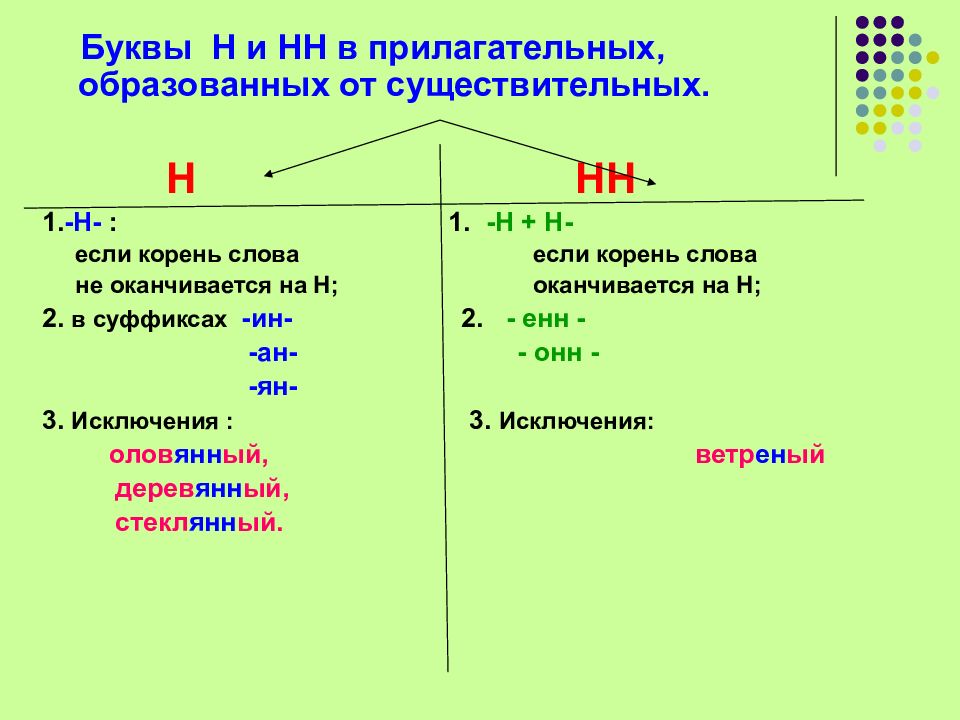
Подготовка и трансформация протопластов.
Приготовление протопластов риса и трансформацию проводили, как описано (52 и 53) с небольшими модификациями. Для приготовления протопластов стебли и оболочки 14-дневных проростков риса TNG67 разрезали на полоски 0,5 мм и инкубировали в растворе фермента [2% целлюлазы RS (Yakult), 1% мацерозима R10 (Yakult), 0,1% Mes, pH 5,6, 0,6 M маннита, 0,1% CaCl 2 и 1% BSA] и подвергали вакуумной инфильтрации (15–20 см рт. Ст.) В течение 15 мин.После вакуумной инфильтрации полоски в растворе фермента осторожно встряхивали на свету в течение примерно 3,5 ч, пока протопласты не перешли в раствор. После переваривания растворы, содержащие протопласты, фильтровали через нейлоновые сетки 40 мкм с последующим центрифугированием при 200 × г в течение 3 минут с качающимся ведром для осаждения протопластов в пробирке с круглым дном. Супернатанты удаляли, осадки протопластов ресуспендировали в растворе W5 [154 мМ NaCl, 125 мМ CaCl 2 , 5 мМ KCl, 5 мМ глюкозы и 2 мМ 4-морфолинэтансульфоновой кислоты (MES), pH 5. 7], и этот шаг повторяли один раз перед инкубацией протопластов и их ресуспендированием на льду в течение не менее 30 мин. Затем раствор W5 удаляли и протопласты ресуспендировали до конечной концентрации 2–5 × 10 5 клеток на миллилитр в растворе MMG (0,6 М маннита, 15 мМ MgCl 2 и 4 мМ MES, pH 5,7. ). Для трансформации протопластов всего приблизительно 4 × 10 5 протопластов в 0,2 мл раствора MMG смешивали с 20 мкг плазмидной ДНК на льду в течение 10 мин.Затем добавляли равный объем (приблизительно 220 мкл) раствора ПЭГ-кальция [40% мас. / Об. ПЭГ 4000 (95904; Sigma-Aldrich), 0,6 М маннита и 0,1 М CaCl 2 ] и смесь перемешивали. инкубируют при комнатной температуре (RT) в течение 20 мин. После инкубации медленно и осторожно перемешивали 3 мл раствора W5, и протопласты осаждали центрифугированием при 200 × g в течение 1 мин с качающимся ведром. После двукратной промывки раствором W5 гранулы осторожно ресуспендировали в 1.5 мл раствора W5 и инкубировали в шестилуночных планшетах, покрытых 1% BSA, при комнатной температуре в течение указанного времени в темноте.
7], и этот шаг повторяли один раз перед инкубацией протопластов и их ресуспендированием на льду в течение не менее 30 мин. Затем раствор W5 удаляли и протопласты ресуспендировали до конечной концентрации 2–5 × 10 5 клеток на миллилитр в растворе MMG (0,6 М маннита, 15 мМ MgCl 2 и 4 мМ MES, pH 5,7. ). Для трансформации протопластов всего приблизительно 4 × 10 5 протопластов в 0,2 мл раствора MMG смешивали с 20 мкг плазмидной ДНК на льду в течение 10 мин.Затем добавляли равный объем (приблизительно 220 мкл) раствора ПЭГ-кальция [40% мас. / Об. ПЭГ 4000 (95904; Sigma-Aldrich), 0,6 М маннита и 0,1 М CaCl 2 ] и смесь перемешивали. инкубируют при комнатной температуре (RT) в течение 20 мин. После инкубации медленно и осторожно перемешивали 3 мл раствора W5, и протопласты осаждали центрифугированием при 200 × g в течение 1 мин с качающимся ведром. После двукратной промывки раствором W5 гранулы осторожно ресуспендировали в 1.5 мл раствора W5 и инкубировали в шестилуночных планшетах, покрытых 1% BSA, при комнатной температуре в течение указанного времени в темноте.
Анализ ChIP-qPCR.
Для обнаружения прямых генов-мишеней SUB1A-1 были выполнены анализы ChIP-qPCR с использованием системы протопластов риса. Сшивание проводили, как описано (54) с небольшими модификациями. Вкратце, конструкции UbiP :: SUB1A-1-Luc и конструкции Ubi :: Luc (в качестве контроля) трансфицировали в протопласты риса TNG67 соответственно. После 4 ч инкубации при комнатной температуре трансфицированные протопласты (1.6 × 10 ( 6 ( клеток) на трансфекцию) собирали центрифугированием при 200 × g в течение 2 минут при комнатной температуре с последующим удалением супернатантов. Собранные протопласты подвергали поперечному сшиванию 1% формальдегидом в 1,5 мл раствора W5 и осторожно перемешивали на роторе (12 об / мин) в течение 10 мин при комнатной температуре. Чтобы погасить реакцию сшивания, добавляли 80 мкл 2М глицина и осторожно перемешивали на роторе (Intelli Mixer ERM-2L; ELMI) при 12 об / мин в течение 5 минут при комнатной температуре с последующим центрифугированием при 1500 × g в течение 5 минут. мин при 4 ° C для удаления супернатанта, и протопласты промывали 1 мл ледяного буфера 1 × PBS (pH 7.4). Экстракцию хроматина, расщепление МНКазой, обработку ультразвуком, иммунопреципитацию, обратное перекрестное связывание, выделение ДНК и количественную ПЦР проводили с использованием набора Pierce Magnetic ChIP (каталожный номер 26157; Thermo Scientific). Эти процедуры следовали протоколу, предоставленному производителем. Комплекс ДНК-белок подвергали иммунной преципитации с помощью антител против люциферазы (sc-74548; Santa Cruz) в концентрации 5 мкг для каждой иммунопреципитации. Затем связанные фрагменты ДНК были обратно высвобождены и амплифицированы с помощью специфической реакции qPCR.Праймеры, используемые в анализе ChIP-qPCR, перечислены в приложении SI, таблица S1.
Анализ стабильности белков in vitro.
Анализ стабильности белков in vitro проводили, как описано ранее (19). Для проведения анализа in vitro была создана модифицированная версия вектора экспрессии pTNT (Invitrogen), pTNT4 × MYC, которая последовательно содержит промотор T7 и промотор SP6, лидерную группу β-глобина, фрагмент ccdB, фрагмент 4 × MYC и терминатор T7. . Сначала pTNT дважды переваривали XhoI и XbaI и элюировали гелем для очистки вектора pTNT с липкими концами.Плазмиду pGWB516 использовали в качестве матрицы для амплификации фрагментов ccdB, несущих сайты XhoI и EcoRV на 5′- и 3′-концах, и 4 × MYC-фрагментов, несущих сайты EcoRV и XbaI на 5′- и 3′-концах, с помощью ПЦР, соответственно. Используемые здесь праймеры перечислены в приложении SI , таблица S2. Затем амплифицированные фрагменты ccdB дважды расщепляли с помощью XhoI и EcoRV, а амплифицированные фрагменты 4 × MYC дважды переваривали с помощью EcoRV и XbaI. После элюции из геля фрагмент ccdB и фрагмент 4 × MYC были объединены в векторы pTNT с липкими концами для создания pTNT4 × MYC.Подробная информация о последовательности pTNT4 × MYC показана в приложении SI, рис. S10. CDS ERF66 и ERF67 были клонированы из кДНК, полученной из погруженной кДНК FR13A, а фрагменты ДНК ERF66 и ERF67 были субклонированы в pTNT4 × MYC с помощью системы шлюза (Invitrogen) для получения C-концевых слияния с MYC, управляемой Промотор Т7. CDS SUB1A-2 был клонирован из кДНК, полученной из погруженной кДНК IR29, и лигирован в модифицированную pTNT3 × HA (19) для получения слияния с C-концевой HA-меткой, управляемой промотором T7.PTNT3 × HA-SUB1A-1 был получен от Gibbs et al. (19). N-концевые мутации включали путем соответствующего изменения последовательностей прямых праймеров ( SI, приложение , таблица S2). Анализы стабильности белка in vitro проводили с использованием системы лизата ретикулоцитов кролика (L4960; Promega) с добавлением 100 мкМ CHX для блокирования трансляции мРНК. Реакции сначала инкубировали в течение 30 минут при 30 ° C, чтобы обеспечить трансляцию белка. По истечении этого 30-минутного периода добавляется CHX для предотвращения дальнейшей трансляции, и сразу же берется образец реакций (т.е., в момент времени 0), а затем следующие образцы были взяты в указанные моменты времени перед смешиванием с красителем, загружающим белок, для прекращения синтеза белка. Равные количества каждой реакции подвергали иммуноблот-анализу против HA / MYC. Все блоты проверяли на равную загрузку путем окрашивания по Понсо.
Вестерн-блот-анализ и антитела.
Экстракцию белка из трансфицированных протопластов риса (4 × 10 5 клеток на трансфекцию) проводили, как описано (55). Экстракцию белка из трансгенного проростка Arabidopsis проводили, как описано (56).Белки, разделенные с помощью SDS / PAGE, переносили в PVDF с использованием ячейки для электрофоретического переноса MiniTrans-Blot (Bio-Rad). Мембраны зондировали первичными антителами со следующими титрами: анти-НА (h4663; Sigma-Aldrich), 1: 1000; анти-MYC (WH0004609M2; Sigma-Aldrich), 1: 1000; антитубулин (T5168; Sigma-Aldrich), 1: 5,000; анти-GUS (G5545; Sigma-Aldrich), 1: 1000; и анти-люцифераза (sc-74548; Santa Cruz), 1: 200. HRP-конъюгированные вторичные антитела против мыши (NEF822001EA; PerkinElmer) / кролика (DC03L; Calbiochem) использовали в титре 1: 3000.Иммуноблоты визуализировали с помощью реагента с усиленной хемилюминесценцией (SuperSignal West Pico; Thermo Scientific). Относительная интенсивность изображения была определена количественно с помощью ImageJ (https://imagej.nih.gov/ij/).
Экспрессия и очистка белков.
pET32a-SUB1A-1 и pET32a-SUB1A-1N были трансформированы в Escherichia coli Rosetta (DE3). Экспрессию рекомбинантного белка индуцировали при OD 0,6 добавлением 1 мМ IPTG при 25 ° C в течение 6 часов и собирали центрифугированием при 4700 × g в течение 30 минут.Клетки ресуспендировали в буфере для лизиса A (50 мМ Hepes, pH 7,5, 400 мМ NaCl и 20 мМ имидазол) с добавлением 10 мкг / мл ДНКазы I, 1 мг / мл лизоцима и 1 мМ PMSF и лизировали обработкой ультразвуком. Лизат центрифугировали при 20000 × g в течение 30 минут, и супернатант загружали в колонку, содержащую смолу NiNTA, предварительно уравновешенную буфером для лизиса. Колонку промывали 20 объемами колонки (CV) буфера для лизиса, а затем 5 объемами промывочного буфера (50 мМ Hepes, pH 7,5, 300 мМ NaCl и 50 мМ имидазол).Белки элюировали элюирующим буфером (50 мМ Hepes, pH 7,5, 100 мМ NaCl, 500 мМ имидазол). После удаления тиоредоксиновой метки путем обработки протеазой TEV в течение ночи при 4 ° C раствор загружали в колонку HiPrep Heparin FF 16/10, предварительно уравновешенную буфером A (50 мМ Hepes, pH 7,5, 10 мМ β-меркаптоэтанол и 5% глицерин). с 100 мМ NaCl. После промывки колонки белок элюировали градиентом 0–100% буфера А с 1 М NaCl в 20 CV. Фракции основного пика, содержащие белок, концентрировали и полировали с использованием колонки ENrich SEC650 с буфером А с 300 мМ NaCl.
Для 15 N-меченого N-конца SUB1A-1, E. coli Rosetta (DE3) культивировали в бульоне Лурия до достижения OD 1,0 и центрифугировали при 4000 × g в течение 20 мин. Затем осадки клеток промывали и ресуспендировали в буфере M9 три раза. Ресуспендированные клетки восстанавливали в среде M9 с [ 15 N] NH 4 Cl в качестве единственного источника азота в течение 1 ч при 37 ° C перед индукцией в течение ночи при 16 ° C путем добавления 0,5 мМ IPTG.
КД-спектрометрия.
Спектры КД в дальней УФ-области регистрировали в диапазоне 204–260 нм при 25 ° C с использованием спектрометра Jasco J-815 (Jasco).Всего 5,5 мкМ SUB1A-1 и 12 мкМ SUB1A-1N (оба в 5 мМ Hepes, pH 7,5 и 100 мМ NaCl) были перенесены в кварцевую кювету диаметром 1 мм перед сбором данных. Все спектры вычитали из буфера и сглаживали с помощью Spectra Analysis (Jasco). Результаты выражаются в виде средней остаточной молярной эллиптичности.
ЯМР-спектроскопия.
Образец ЯМР в пробирке Shigemi с OD 4 мм содержал 200 мкл, pH 7,0, водный буферный раствор (90% H 2 O / 10% D 2 O) с 0.14 мМ 15 N-меченный белок SUB1A-1N, 150 мМ NaCl, 25 мМ фосфат калия, 1 мМ NaN 3 и 0,1 мМ 4,4-диметил-4-силапентан-1-сульфоновая кислота в качестве внутреннего химического вещества стандарт смены. Все данные ЯМР собирали при 298 К на ЯМР-спектрометре Bruker 800 МГц (AV800), оборудованном криогенным зондом TXI. Двумерные спектры 1 H, 15 N-HSQC были получены по схеме BEST (57). Параметры регистрации для BEST 2D 1 H, 15 N-HSQC были следующими: центр импульсов селективности протонов N-H на 8.5 ppm, задержка между сканированием 0,2 с, 512 сканирований на приращение, 256 приращений в сумме измерения 15 N. Обработанные растворителем 2D 1 H, 15 Данные N-HSQC были собраны с использованием схемы химического обмена CLEAN с фазовой модуляцией (58) с обменным перемешиванием 100 мс, 360 сканирований на приращение и 128 приращений в 15 Размер Н. Данные ЯМР обрабатывали с помощью программного обеспечения Topspin (Bruker).
iTC Binding Assays.
Эксперимент iTC проводили на калориметре ITC200 (MicroCal) при 25 ° C.Буфер для измерения состоял из 50 мМ Hepes, 100 мМ NaCl и 1 мМ DTT, pH 8,0. Шприц для инъекций (40 мкл) заполняли 1 мМ N-концом SUB1A-1, а в ячейку для образца загружали 200 мкл 100 мкМ С-конца SUB1A-1. За первой инъекцией (0,3 мкл) N-домена SUB1A-1 следовали 13 инъекций по 3 мкл при скорости перемешивания 1000 об / мин. Значение титрования первой инъекции не использовалось при анализе данных. Наилучшее соответствие изотерм привязки было получено путем вычитания насыщенного интеграла сигнала из последней точки в качестве эталона.Однако взаимодействие относительно слабое, поэтому термодинамические параметры (N, ΔH, K A ) не могут быть точно оценены. Все данные были нанесены на график и проанализированы с помощью программного обеспечения Microcal Origin.
Подробные сведения об экстракции РНК и qRT-PCR, RNA-seq и анализе данных, конструировании плазмиды, анализе активации транс , EMSA и дрожжевом двухгибридном анализе описаны в приложении SI , SI Материалы и методы . Все праймеры для qRT-PCR и клонирования кДНК, а также зонды для EMSA перечислены в приложении SI, приложение , таблицы S1 – S3.Благодарности тепличное ядро Academia Sinica за техническую поддержку. Мы признательны за использование инструментов в основной лаборатории биофизики Центра научных инструментов Академии Синица. Мы благодарим Academia Sinica High-Field NMR Center (HFNMRC) за техническую поддержку; HFNMRC финансируется проектом Academia Sinica Core Facility и Innovative Instrument Project (AS-CFII-108-112).Работа выполнена при поддержке Academia Sinica и Министерства науки и технологий, грант MOST 107-2311-B-001-002 (для M.-CH), грант Taiwan Protein Project, AS-KPQ-105-TPP (для M.-CH). , Грант Совета по биотехнологиям и биологическим научным исследованиям BB / M020568 / 1 (для DJG) и Стартовый грант Европейского исследовательского совета 715441-GasPlaNt (для DJG).
Сноски
Вклад авторов: C.-C.L., S.-J.C., M.-C.H. и M.-C.S. спланированное исследование; C.-C.L., W.-C.C., H.-Y.H., M.-YC, Y.-RL, Y.-LW, H.-AY, HH, C.-SL, F.-HW, H.-CJ, Y.-HH, DI, W.- JW, J.-LW, DJG и M.-CH проведенное исследование; Y.-T.C., C.-S.L., F.-H.W., H.-C.J., Y.-H.H., D.I., W.-J.W., J.-L.W. и M.-C.H. внесены новые реагенты / аналитические инструменты; C.-CL, Y.-TC, W.-CC, H.-YH, M.-YC, Y.-RL, Y.-LW, H.-AY, HH, S.-JC, DJG, M .-CH и М.-CS проанализированные данные; и C.-C.L., D.J.G., M.-C.H. и M.-C.S. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Размещение данных: данные, представленные в этом документе, были депонированы в базе данных Национального центра биотехнологической информации BioProject, https://www.ncbi.nlm.nih.gov/bioproject (инвентарный номер PRJNA512592).
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1818507116/-/DCSupplemental.
Пикорноподобный вирус подавляет путь правила N-конца, подавляя апоптоз
Основные комментарии:
1) Чтобы определить метаболическую стабильность белков DIAP1 и NTAN1, авторы измерили их стабильные уровни, используя только анализ иммуноблоттинга.Результаты иммуноблоттинга недостаточны для того, чтобы сделать выводы о деградации белка, и должны быть подтверждены с помощью анализов деградации in vivo, таких как анализ с отслеживанием импульсов. Если импульсный анализ технически невозможен, альтернативой являются анализы разложения циклогексимида.
Мы искренне признательны и согласны с этим ценным комментарием. Следуя этому предложению, мы провели анализы деградации CHX для DIAP1, NTAN1 и NTAN1-4KA (мутант со всеми 4 остатками лизина, мутированными в аланин).Данные анализов деградации CHX показывают, что вирусная инфекция способствовала деградации этих белков (новый рисунок 3 — рисунок, дополнение 1; новый рисунок 5E-H; новый рисунок 6D-H; подраздел «Вирусная инфекция способствует накоплению расщепленного DIAP1 в клетках. », Первый абзац; подраздел« Вирусная инфекция способствует истощению NTAN1 на ранней стадии инфекции », второй абзац и подраздел« Вирусная инфекция способствует деградации NTAN1 через протеасомный путь », второй абзац).
2) Авторы утверждают, что индуцированная вирусом деградация NTAN1 опосредуется протеасомой без убиквитинирования. В общем, такой отрицательный вывод с убиквитинированием требует более строгой проверки. Например, во втором абзаце подраздела «Вирусная инфекция способствует деградации NTAN1 через протеасомный путь» (рис. 6C), авторы утверждают, что мутация всех четырех остатков лизина не ингибирует деградацию NTAN, и предположили, что индуцированная вирусом деградация NTAN1 не зависит от убиквитинирования.Этот иммуноблоттинг-анализ низкого качества. Распад NTAN1-4KA следует сравнивать с NTAN1 дикого типа. Кроме того, результаты должны быть подтверждены с помощью анализа последовательности импульсов или, по крайней мере, анализа разложения циклогексимида. Рецензенту кажется, что NTAN1-4KA значительно стабилизировался на 9 л.с. по сравнению со всеми четырьмя одиночными мутантами по лизину (Рисунок 6 — рисунок в приложении 2).
Искренне благодарим Вас за ценные комментарии. Следуя этому предложению, мы снова провели иммуноблоттинг на рисунке 6C с несколькими повторами и заменили его на показатель лучшего качества в пересмотренной версии (новый рисунок 6C).
Как мы ответили на главный комментарий № 1, мы сравнили распад NTAN1-4KA с NTAN1 WT, используя анализ деградации CHX. Наши данные показывают, что вирусная инфекция может способствовать деградации как мутанта 4KA, так и WT NTAN1 (новый рисунок 6D-H; подраздел «Вирусная инфекция способствует деградации NTAN1 через протеасомный путь», второй абзац), хотя NTAN1-4KA более стабилен. чем WT NTAN1 в отсутствие инфекции DCV (новые рисунки 6D, F и H). Кроме того, на основании наших данных и количественной оценки на Фигуре 6C, вирусная инфекция вызвала деградацию NTAN1-4KA через 9 часов.p.i, аналогично WT и другим мутантам. Фактически, есть два способа деградировать NTAN1 (Ub-зависимый и –независимый) одновременно. Мутация NTAN1-4KA может полностью блокировать путь Ub-зависимой деградации, в то время как единичная мутация лизина не может полностью блокировать этот путь. Таким образом, ожидается, что NTAN1-4KA более стабилен, чем другие мутанты по одиночному лизину в присутствии или отсутствии вирусной инфекции.
3) Авторы утверждают, что вирусная инфекция вызывает деградацию NTAN1, но не исследовали скорость распада эндогенного NTAN1 в нормально растущих клетках.Необходимо сравнить скорости деградации NTAN1 в двух условиях с вирусной инфекцией или без нее, используя анализы деградации in vivo.
Мы согласны с этим комментарием. Следуя этому предположению, мы исследовали скорость деградации эндогенного NTAN1 в нормально растущих клетках как в присутствии, так и в отсутствие вирусной инфекции, используя анализ деградации CHX. И наши данные показывают, что вирусная инфекция может вызывать эндогенную деградацию NTAN1 (новый рисунок 5E-H; подраздел «Вирусная инфекция способствует истощению NTAN1 на ранней стадии инфекции», второй абзац).
4) Одним из основных недостатков этого исследования является отсутствие какого-либо механистического понимания того, какие вирусные компоненты участвуют в нацеливании на NTAN1. Кажется, довольно просто рассмотреть эффекты белков, введенных вирусом, и выполнить хотя бы базовые тесты, чтобы увидеть, как они могут потенциально повлиять на стабильность NTAN1. Авторы не предлагают никакого понимания этого, даже не обсуждают возможности. Хотя я понимаю, что подробные эксперименты в этом направлении могут быть трудными, я полагаю, что документ может быть значительно усилен, по крайней мере, обсуждением или моделью.
Мы искренне признательны за этот ценный комментарий. Действительно, мы изо всех сил старались выяснить точный механизм, с помощью которого инфекция DCV способствует деградации NTAN1. С этой целью мы попытались экспрессировать отдельные белки DCV в культивируемых клетках, чтобы изучить, как они могут потенциально влиять на стабильность NTAN1, как также предположил автор обзора. Однако из-за ограниченности современных знаний точные границы этих отдельных вирусных белков в полипротеинах DCV неясны (геном DCV содержит две ORF, которые кодируют два полипротеина, которые впоследствии расщепляются DCV 3C-подобной протеазой на отдельные белки).На основе анализа последовательностей и сопоставления с другими пикорнавирусами и пикорна-подобными вирусами мы предсказали последовательности этих белков DCV (1A, Helicase, RdRP, 3C-like, VP1, VP2, VP3 и VP4) и клонировали их в векторы экспрессии. К сожалению, большинство этих вирусных белков было чрезвычайно трудно экспрессировать в клетках S2 (это явление не является необычным для экспрессии многих других вирусных белков). Мы успешно экспрессировали DCV 3C-подобную протеазу (3CL) в клетках S2, которая является идеальным кандидатом, поскольку протеаза пикорнавируса 3C, как сообщается, расщепляет многие белки-хозяева.Однако мы не наблюдали какого-либо влияния 3CL на стабильность NTAN1 (данные не показаны в рукописи). Следуя предложению этого рецензента, мы обсудили возможные механизмы, которые могут вызвать деградацию NTAN1 в исправленной рукописи (Обсуждение, пятый абзац).
5) Вестерн-блоттинг во всех панелях должен быть количественно определен с достаточным количеством повторов. Для многих панелей показан только один повтор, поэтому неясно даже, сколько экспериментов было проведено. Очень важно оценивать данные в трех экземплярах, учитывая вариабельность самого метода.
Искренне благодарим за комментарий. Все анализы вестерн-блоттинга были независимо повторены не менее трех раз. Следуя этому предложению, мы количественно оценили данные вестерн-блоттинга с помощью денситометрии на исправленных рисунках, а также описали метод в пересмотренном разделе «Материалы и методы» (подраздел «Вестерн-блоттинг, иммунопреципитация (IP) и антитела», первый абзац).
6) Во многих панелях наблюдается последовательное снижение уровня тубулина с течением времени после инфицирования.Похоже, что вирус воздействует на микротрубочки, что, вероятно, неудивительно, но это поднимает вопросы контроля загрузки. Потенциальным контролем загрузки может быть общая загрузка белка, что также можно проиллюстрировать окрашиванием кумасси синим. Существуют также другие часто используемые элементы управления загрузкой, такие как GAPDH или актин, которые, конечно, также могут изменяться в зависимости от заражения вирусом. Это необходимо решить.
Мы искренне признательны и согласны с этим ценным комментарием.Мы постоянно наблюдали, что вирусная инфекция может вызывать снижение уровня тубулина после 24-часовой инфекции. Следуя этому комментарию, мы использовали общие белковые нагрузки в качестве контроля загрузки с помощью окрашивания кумасси синим (новый рисунок 3A) и соответствующим образом изменили раздел «Материалы и методы» (подраздел «Вестерн-блоттинг, иммунопреципитация (IP) и антитела», первый абзац). . Кроме того, по нашим собственным наблюдениям, GAPDH или актин нельзя использовать в качестве хорошего контроля нагрузки, поскольку их коммерческие антитела обычно распознают множество шумовых полос в вестерн-блотах экстрактов клеток Drosophila S2.
7) Аналогичным образом количественная оценка уровней трансфицированного белка должна сопровождаться данными qPCR, показывающими уровни трансфекции, поскольку они могут значительно повлиять на результат между культурами. «Быстрый и грязный» способ — разделить культуру поровну после трансфекции в лунки / чашки, используемые для сбора временных точек. Я не нашел никаких экспериментальных подробностей об этом в рукописи.
Мы согласны с этим комментарием.Собственно, в наших экспериментах мы уже использовали «быстрый и грязный» способ, предложенный рецензентами. Вкратце, мы сначала трансфицировали клетки в 100-мм культуральной чашке. После 24-36 часов трансфекции трансфицированные клетки делили на шесть или восемь лунок 6-луночного планшета и культивировали еще 6 часов. Затем клетки подвергали вирусной инфекции или другим обработкам в соответствии с экспериментальными требованиями. Исходя из этой процедуры, уровень трансфекции или эффективность в разных лунках должны быть одинаковыми.Процедуры эксперимента подробно описаны в отредактированной рукописи (подраздел «Трансфекция»).
Кроме того, следуя предложению авторов обзора, мы количественно определили уровни трансфекции с помощью кПЦР, и наши данные показывают, что при использовании вышеуказанного метода уровни трансфекции в разных образцах были схожими (новый рисунок 3 — приложение к рисунку 2, подраздел «Вирусная инфекция»). способствует накоплению расщепленного DIAP1 в клетках », третий абзац).
8) На фиг. 7 дополнительный NTAN1 усиливает вирус-индуцированный апоптоз и снижает репликацию вируса в клетке.Как насчет клеток с нокдауном NTAN1? На Фигуре 4C клетки, разрушенные дцРНК ATE1 и NTAN1, демонстрируют накопление расщепленной формы DAIP1. Важно исследовать уровень каспаз или апоптотическое состояние и гРНК DCV в клетках с нокдауном NTAN1 при заражении DCV.
Искренне благодарим рецензентов за комментарий. Следуя этому предложению, мы изучили эффект нокдауна NTAN1, и наши данные показывают, что его нокдаун ингибирует апоптоз, индуцированный вирусом, а также усиливает репликацию DCV (новый рисунок 7 — добавление к рисунку 1, подраздел «Индуцированная вирусом деградация NTAN1 ингибирует апоптоз и дает преимущества. вирусная репликация »).
9) Механизм противовирусной функции апоптоза не ясен. Повышенный уровень гРНК DCV в клетках можно объяснить многими причинами. Репликация вирусной РНК была усилена в клетках с помощью антиапоптотического DIAPI, путь образования вируса был заблокирован, так что вирусная РНК накапливалась в клетках, или проникновение вируса было более эффективным, чем в других группах, и т. Д. Реплицированный геном РНК упакован и упакован заново. частицы должны попадать в культуральные жидкости. Одним из возможных экспериментов является анализ вирусных частиц в культивируемых жидкостях либо RT-qPCR, либо любыми доступными методами для решения проблемы повышенного уровня DCV gRNA, что означает более жизнеспособный DCV.
Мы благодарны за этот комментарий. Следуя этому предложению, мы определили уровни вирусных частиц в культуральных жидкостях с помощью КПЦР. Наши данные показывают, что сверхэкспрессия DIAP1 или подавление каспаз не приводит к снижению уровня вирусов в культивируемых жидкостях (новые рисунки 2C и F, подраздел «Ингибирование апоптоза усиливает репликацию вирусов в клетках и взрослых мухах», первый абзац), подтверждая, что репликация вирусной РНК была усилена в клетках, ингибированных апоптозом.
https://doi.org/10.7554/eLife.30590.025
(n + 1) Правило — Chemistry LibreTexts
Правило (n + 1), эмпирическое правило, используемое для прогнозирования множественности и, в сочетании с треугольником Паскаля, структуры пиков в 1 H и 13 C Спектры ЯМР заявляют, что если данное ядро связано (см. Спиновое взаимодействие) с числом n ядер, которые эквивалентны (см. Эквивалентные лиганды), кратность пика равна n + 1. например. 1:
Три ядра водорода в 1, \ (H_a \), \ (H_b \) и \ (H_c \) эквивалентны.Таким образом, в спектре ЯМР 1H 1 \ (H_a \) s только один пик. \ (H_a \), \ (H_b \) и \ (H_c \) не связаны ни с какими ядрами водорода. Таким образом, для \ (H_a \), \ (H_b \) и \ (H_c \), n = 0; (n + 1) = (0 + 1) = 1. Кратность пика \ (H_a \), \ (H_b \) и \ (H_c \) равна единице. Пик \ (H_a \) — одна линия; это синглет. например. 2:
В 2:
есть два набора эквивалентных ядер водорода.
- Набор 1: \ (H_a \)
- Набор 2: \ (H_b \), \ (H_c \)
Таким образом, в спектре ЯМР 1H 2 \ (H_a \) два пика, один связан с \ (H_a \), а другой — с \ (H_b \) и \ (H_c \).
Пик \ (H_a \): есть два ближайших к \ (H_a \) атома водорода: \ (H_b \) и \ (H_c \). \ (H_b \) и \ (H_c \) эквивалентны друг другу, но не \ (H_a \). Таким образом, для \ (H_a \) n = 2; (n + 1) = (2 + 1) = 3. Кратность пика \ (H_a \) равна трем. Пик \ (H_a \) — три линии; из треугольника Паскаля это тройка.
Пик \ (H_b \) и \ (H_c \): есть только один вицинальный водород для \ (H_b \) и \ (H_c \): \ (H_a \). \ (H_a \) не эквивалентно \ (H_b \) и \ (H_c \).Таким образом, для \ (H_b \) и \ (H_c \) n = 1; (n + 1) = (1 + 1) = 2. Кратность пика \ (H_b \) и \ (H_c \) равна двум. Пик \ (H_a \) — две линии из треугольника Паскаля, это дублет.
Чтобы определить кратность пика ядра, связанного с более чем одним набором эквивалентных ядер, примените правило (n + 1) независимо друг к другу.
например:
В 3:
есть три набора эквивалентных ядер водорода.
- Набор 1: \ (H_a \)
- Набор 2: \ (H_b \)
- Набор 3: \ (H_c \)
пик \ (H_a \):
кратность пика \ (H_a = 2 \ times 2 = 4 \).Чтобы определить схему расщепления пика \ (H_a \), используйте треугольник Паскаля, основываясь на наблюдении, что для алкенильных водородов \ (J_ {cis}> J_ {gem} \).
Пик \ (H_a \) — это дуплет дублета.
пик \ (H_b \):
кратность пика \ (H_b = 2 \ times 2 = 4 \). Чтобы определить схему расщепления пика \ (H_b \), используйте треугольник Паскаля, основываясь на наблюдении, что для алкенильных водородов \ (J_ {trans}> J_ {gem} \).
Пик \ (H_b \) — дублет дублета.
пик \ (H_c \):
кратность пика \ (H_c = 2 \ times 2 = 4 \). Чтобы определить схему расщепления пика \ (H_c \), используйте треугольник Паскаля, основываясь на наблюдении, что для алкенильных водородов \ (J_ {trans}> J_ {cis} \).
Пик \ (H_c \) — дублет дублета.
Авторы и авторство
.